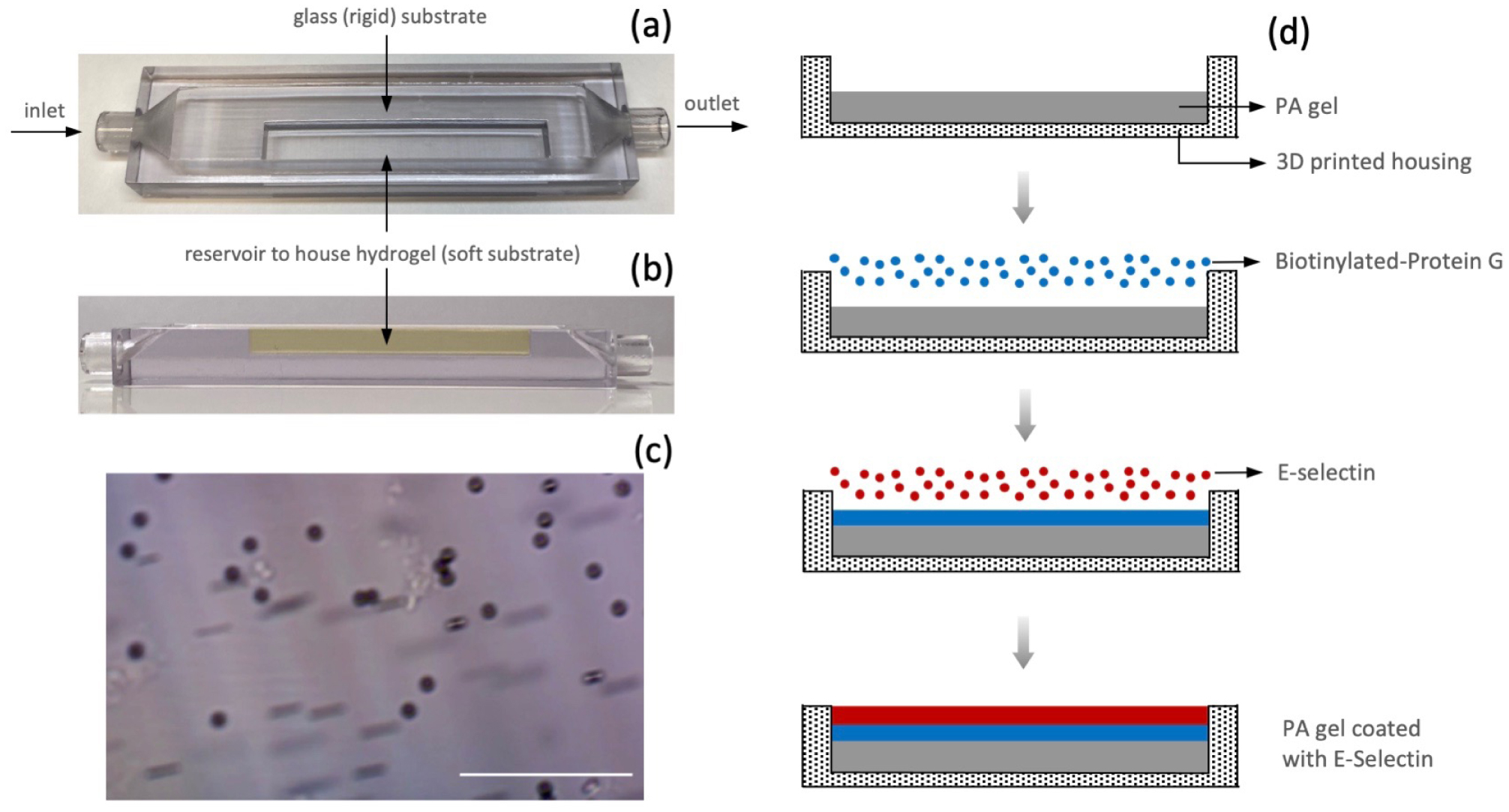
Atherosclerosis is a chronic inflammatory disease that results in the formation of lipid-rich lesions and stiffening of arterial walls. An increasing body of evidence suggests that nearly all members of the leukocyte family accumulate within atherosclerosis-prone arteries and participate in various stages of disease progression. Recently, it has been proposed that progressive changes of the elastic modulus of the arterial wall during plaque development may directly influence the kinematics of leukocyte rolling. In the present study, we propose that rigidity sensing of rolling leukocytes may occur spontaneously due to the stiffness-dependent elastic instability of reversible bonds between rolling leukocytes and the arterial walls. This effect is mechanistic in nature and operates independently of cell biochemical signaling. To partially test this hypothesis, we measured the rolling velocities of functionalized microparticles, comparable in size to leukocytes, interacting with E-selectin coated substrates of controlled stiffness. The kinematic analysis of the particles' motion reveals a larger rolling velocity on softer substrates, aligning with previous reports regarding monocytes. A simple kinetic model for a cluster of reversible bonds formed between a cell and the underlying substrate demonstrates that the critical forces needed for bond disassembly decrease as substrate stiffness decreases. Consequently, bonds are more likely to break on softer substrates, resulting in enhanced cell mobility.
Citation: Alireza Sarvestami, Madeline Smith, Arsha Moorthy, Patrick Kho, Lauren Talbo, Chamaree de Silva. Rigidity sensing by blood-borne leukocytes: Is it independent of internal signaling?[J]. AIMS Biophysics, 2024, 11(1): 18-30. doi: 10.3934/biophy.2024002
Atherosclerosis is a chronic inflammatory disease that results in the formation of lipid-rich lesions and stiffening of arterial walls. An increasing body of evidence suggests that nearly all members of the leukocyte family accumulate within atherosclerosis-prone arteries and participate in various stages of disease progression. Recently, it has been proposed that progressive changes of the elastic modulus of the arterial wall during plaque development may directly influence the kinematics of leukocyte rolling. In the present study, we propose that rigidity sensing of rolling leukocytes may occur spontaneously due to the stiffness-dependent elastic instability of reversible bonds between rolling leukocytes and the arterial walls. This effect is mechanistic in nature and operates independently of cell biochemical signaling. To partially test this hypothesis, we measured the rolling velocities of functionalized microparticles, comparable in size to leukocytes, interacting with E-selectin coated substrates of controlled stiffness. The kinematic analysis of the particles' motion reveals a larger rolling velocity on softer substrates, aligning with previous reports regarding monocytes. A simple kinetic model for a cluster of reversible bonds formed between a cell and the underlying substrate demonstrates that the critical forces needed for bond disassembly decrease as substrate stiffness decreases. Consequently, bonds are more likely to break on softer substrates, resulting in enhanced cell mobility.

| [1] | Ley K (2013) Physiology of Inflammation. New York: Springer. https://doi.org/10.1007/978-1-4614-7512-5 |
| [2] |
McEver RP (1994) Selectins. Curr Opin Immunol 6: 75-84. https://doi.org/10.1016/0952-7915(94)90037-X 
|
| [3] |
Patel KD, Cuvelier SL, Wiehler S (2002) Selectins: critical mediators of leukocyte recruitment. Semin Immunol 14: 73-81. https://doi.org/10.1006/smim.2001.0344
|
| [4] |
Varki A (1994) Selectin ligands. Proc Natl Acad Sci 91: 7390-7397. https://doi.org/10.1073/pnas.91.16.7390
|
| [5] |
Rosen SD, Bertozzi CR (1994) The selectins and their ligands. Curr Opin Cell Biol 6: 663-673. https://doi.org/10.1016/0955-0674(94)90092-2
|
| [6] |
Zarbockn A, Müller H, Kuwano Y, et al. (2009) Psgl-1-dependent myeloid leukocyte activation. J Leukocyte Biol 86: 1119-1124. https://doi.org/10.1189/jlb.0209117
|
| [7] |
Carlow DA, Gossens K, Naus S, et al. (2009) Psgl-1 function in immunity and steady state homeostasis. Immunol Rev 230: 75-96. https://doi.org/10.1111/j.1600-065X.2009.00797.x
|
| [8] |
Zarbock A, Ley K, McEver RP, et al. (2011) Leukocyte ligands for endothelial selectins: specialized glycoconjugates that mediate rolling and signaling under flow. Blood 118: 6743-675. https://doi.org/10.1182/blood-2011-07-343566
|
| [9] |
McEver RP, Cummings RD (1997) Perspectives series: cell adhesion in vascular biology. role of psgl-1 binding to selectins in leukocyte recruitment. J Clin Invest 100: 485-491. https://doi.org/10.1172/jci119556
|
| [10] |
Lo CM, Wang HB, Dembo M, et al. (2000) Cell movement is guided by the rigidity of the substrate. Biophys J 79: 144-152. https://doi.org/10.1016/S0006-3495(00)76279-5
|
| [11] |
Yeung T, Georges PC, Flanagan LA, et al. (2005) Effects of substrate stiffness on cell morphology, cytoskeletal structure, and adhesion. Cell Motil Cytoskel 60: 24-34. https://doi.org/10.1002/cm.20041
|
| [12] |
Giannone G, Sheetz MP (2006) Substrate rigidity and force define form through tyrosine phosphatase and kinase pathways. Trends Cell Biol 16: 213-223. https://doi.org/10.1016/j.tcb.2006.02.005
|
| [13] |
Sheetz M (2019) A tale of two states: normal and transformed, with and without rigidity sensing. Annu Rev Cell Dev Biol 35: 169-190. https://doi.org/10.1146/annurev-cellbio-100818-125227
|
| [14] |
Eriksson EE (2003) Leukocyte recruitment to atherosclerotic lesions, a complex web of dynamic cellular and molecular interactions. Curr Drug Targets Cardiovasc Hematol Disord 3: 309-325. https://doi.org/10.2174/1568006033481357
|
| [15] |
Libby P, Ridker PM, Maseri A (2002) Inflammation and atherosclerosis. Circulation 105: 1135-1143. https://doi.org/10.1146/annurev.pathol.1.110304.100100
|
| [16] |
Sharif AR, Shadpour MT, Avolio A (2019) Progressive changes of elastic moduli of arterial wall and atherosclerotic plaque components during plaque development in human coronary arteries. Med Biol Eng Comput 57: 731-740. https://doi.org/10.1007/s11517-018-1910-4
|
| [17] |
MacKay JL, Hammer DA (2016) Stiff substrates enhance monocytic cell capture through e-selectin but not p-selectin. Integr Biol 8: 62-72. https://doi.org/10.1039/c5ib00199d
|
| [18] | Tse JR, Engler AJ (2010) Preparation of hydrogel substrates with tunable mechanical properties. Curr protoc Vell Biol 47: 10-16. https://doi.org/10.1002/0471143030.cb1016s47 |
| [19] |
Farzi B, Young D, Scrimgeour J, et al. (2019) Mechanical properties of p-selectin psgl-1 bonds. Colloid Surface B 173: 529-538. https://doi.org/10.1016/j.colsurfb.2018.10.017
|
| [20] | Chester B, de Silva C, Judd M, et al. (2022) Capstone project–design of a novel flow chamber to study the effects of vascular stiffness on migration of blood-borne cells. In Proceeding of the American Society for Engineering Education . Charleston, SC: . https://sites.asee.org/se/wp-content/uploads/sites/56/2022/03/2022ASEESE75.pdf |
| [21] |
Athanasiou K, Natoli R (2009) Introduction to Continuum Biomechanics. Springer Cham. https://doi.org/10.1007/978-3-031-01626-4
|
| [22] |
Famularo J, Happel J (1965) Sedimentation of dilute suspensions in creeping motion. AIChE J 11: 981-988. https://doi.org/10.1002/aic.690110608
|
| [23] |
Slack SM, Cui Y, Turitto VT (1993) The effects of flow on blood coagulation and thrombosis. Thromb Haemost 70: 129-134. https://doi.org/10.1055/s-0038-1646173
|
| [24] |
Krasik EF, Hammer DA (2004) A semianalytic model of leukocyte rolling. Biophys J 87: 2919-2930. https://doi.org/10.1529/biophysj.104.039693
|
| [25] |
Kuo SC, Hammer DA, Lauffenburger DA (1997) Simulation of detachment of specifically bound particles from surfaces by shear flow. Biophys J 73: 517-531. https://doi.org/10.1016/s0006-3495(97)78090-1
|
| [26] |
Dembo M, Torney DC, Saxman K, et al. (1988) The reaction-limited kinetics of membrane-to-surface adhesion and detachment. Proc R Soc London B Biol Sci 234: 55-83. https://doi.org/10.1098/rspb.1988.0038
|
| [27] | Johnson KL, Johnson KL (1987) Contact Mechanics. Cambridge university press. https://doi.org/10.1017/CBO9781139171731 |
| [28] | Walcott S, Sun SX (2010) A mechanical model of actin stress fiber formation and substrate elasticity sensing in adherent cells. Biophys J 107: 7757-7762. https://doi.org/10.1073/pnas.0912739107 |
| [29] |
Hammer DA, Apte SM (1992) Simulation of cell rolling and adhesion on surfaces in shear flow: general results and analysis of selectin-mediated neutrophil adhesion. Biophys J 63: 35-57. https://doi.org/10.1016/s0006-3495(92)81577-1
|
| [30] |
Huang JY, Peng XL, Xiong CY, et al. (2011) Influence of substrate stiffness on cell–substrate interfacial adhesion and spreading: a mechano-chemical coupling model. J Colloid Interf Sci 355: 503-508. https://doi.org/10.1016/j.jcis.2010.12.055
|
| [31] |
Dimitriadis EK, Horkay F, Maresca J, et al. (2002) Determination of elastic moduli of thin layers of soft material using the atomic force microscope. Biophys J 82: 2798-2810. https://doi.org/10.1016/S0006-3495(02)75620-8
|
| [32] |
Chang KC, Hammer DA (1996) Influence of direction and type of applied force on the detachment of macromolecularly-bound particles from surfaces. Langmuir 12: 2271-2282. https://doi.org/10.1021/la950690y
|
| [33] |
Zhao YH, Chien S, Skalak R (1995) A stochastic model of leukocyte rolling. Biophys J 69: 1309-1320. https://doi.org/10.1016/s0006-3495(95)79998-2
|





Figures(6) / Tables(1)

Alireza Sarvestami, Madeline Smith, Arsha Moorthy, Patrick Kho, Lauren Talbo, Chamaree de Silva. Rigidity sensing by blood-borne leukocytes: Is it independent of internal signaling?[J]. AIMS Biophysics, 2024, 11(1): 18-30. doi: 10.3934/biophy.2024002







 DownLoad:
DownLoad: