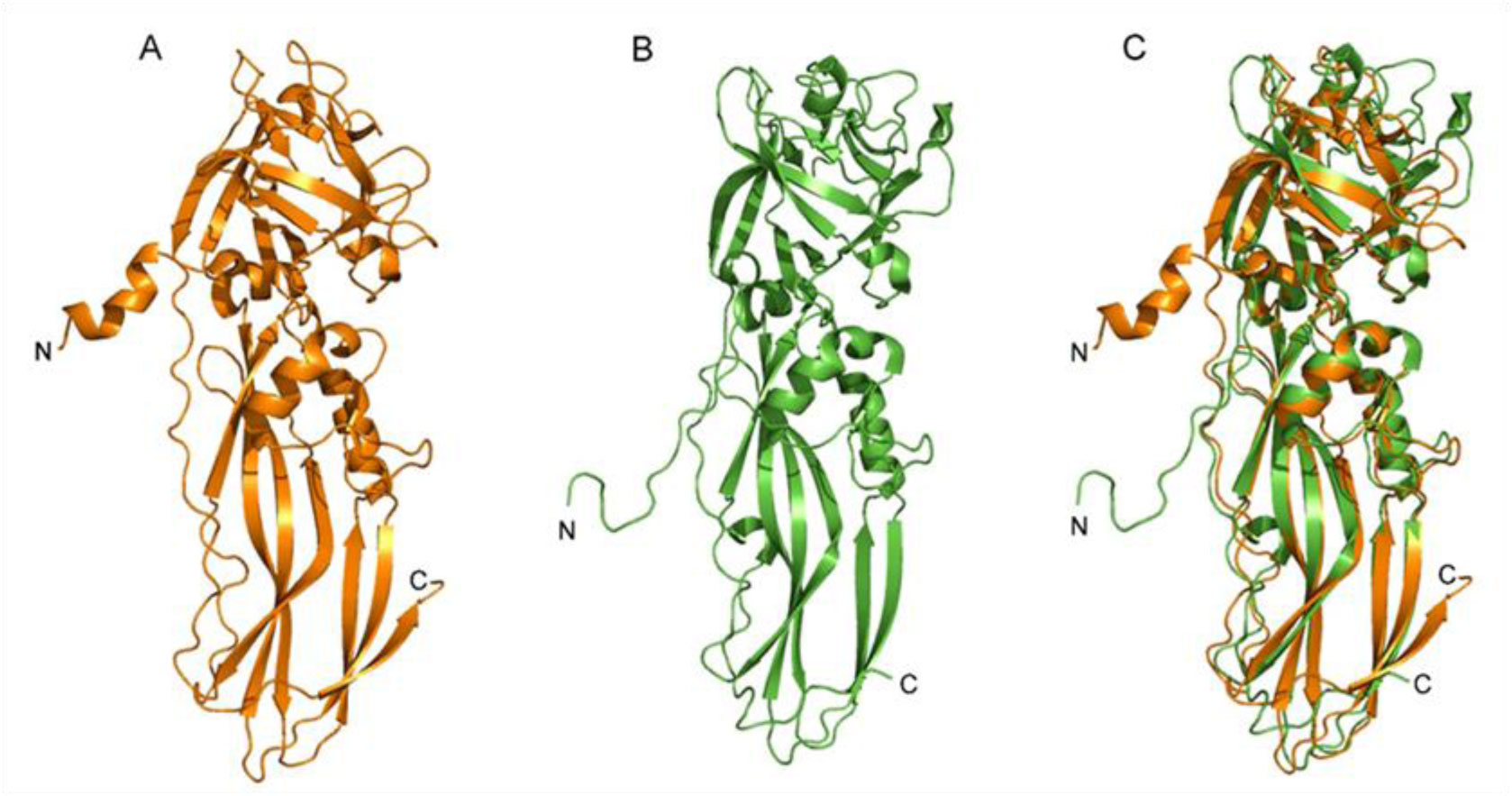
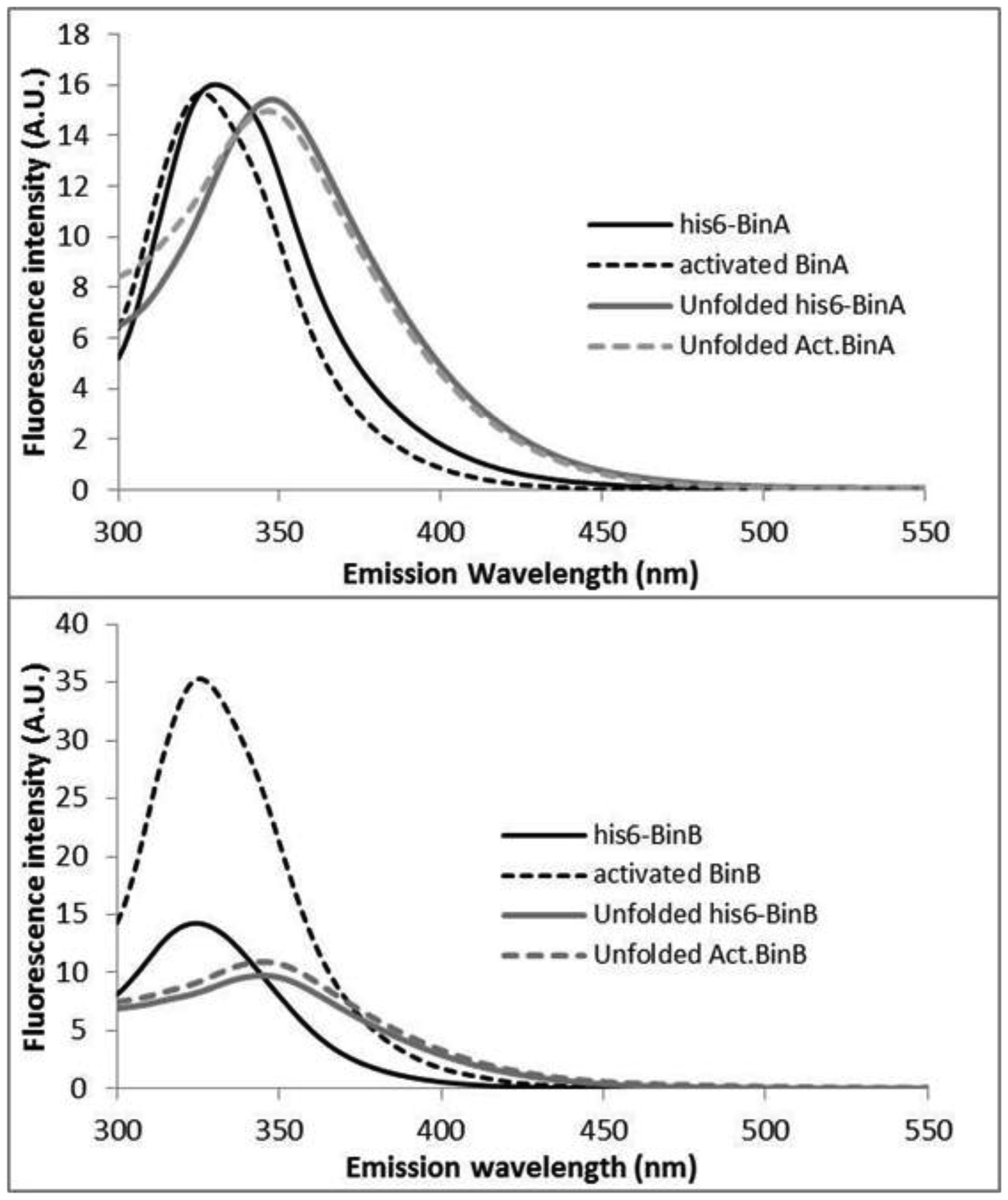
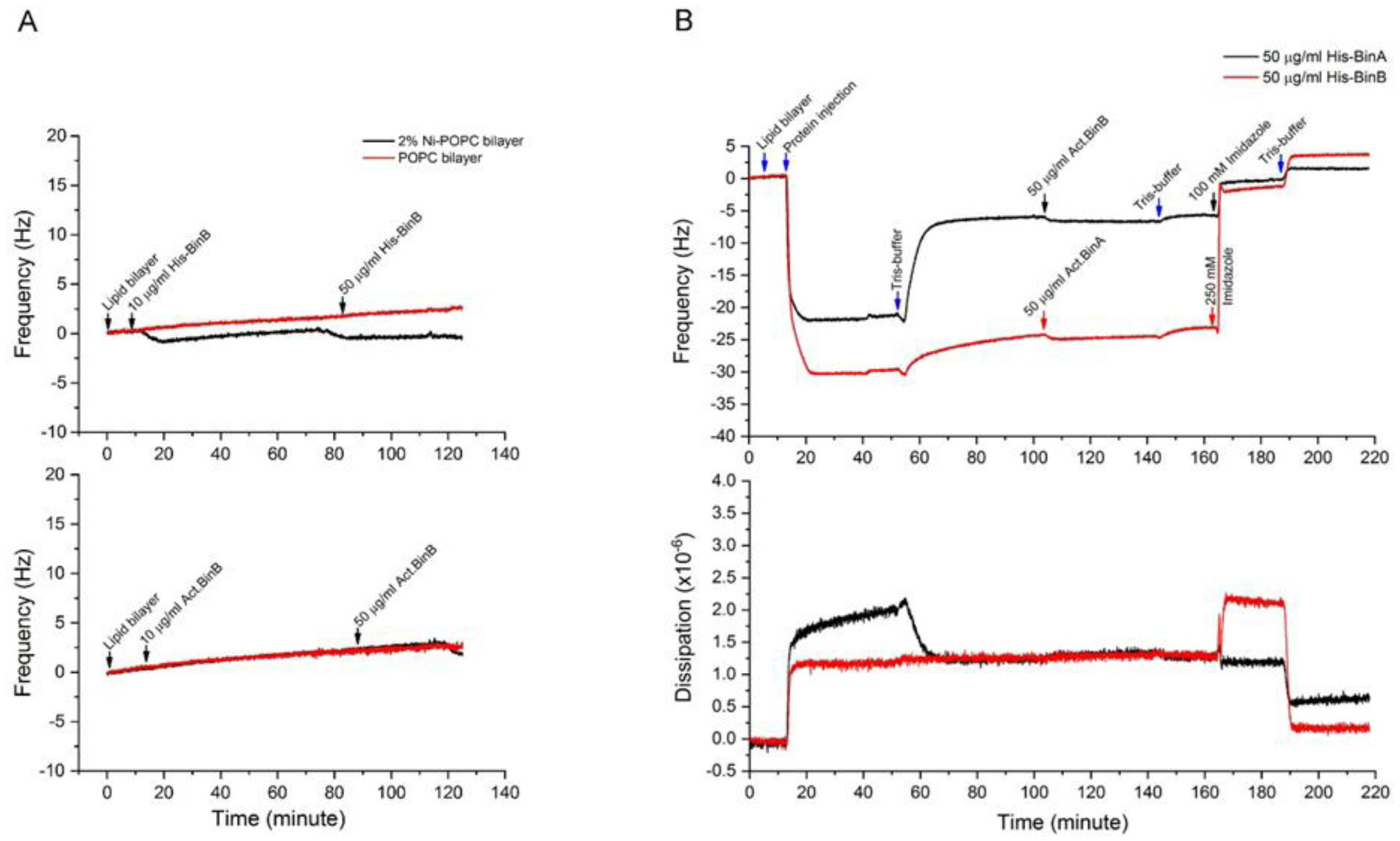
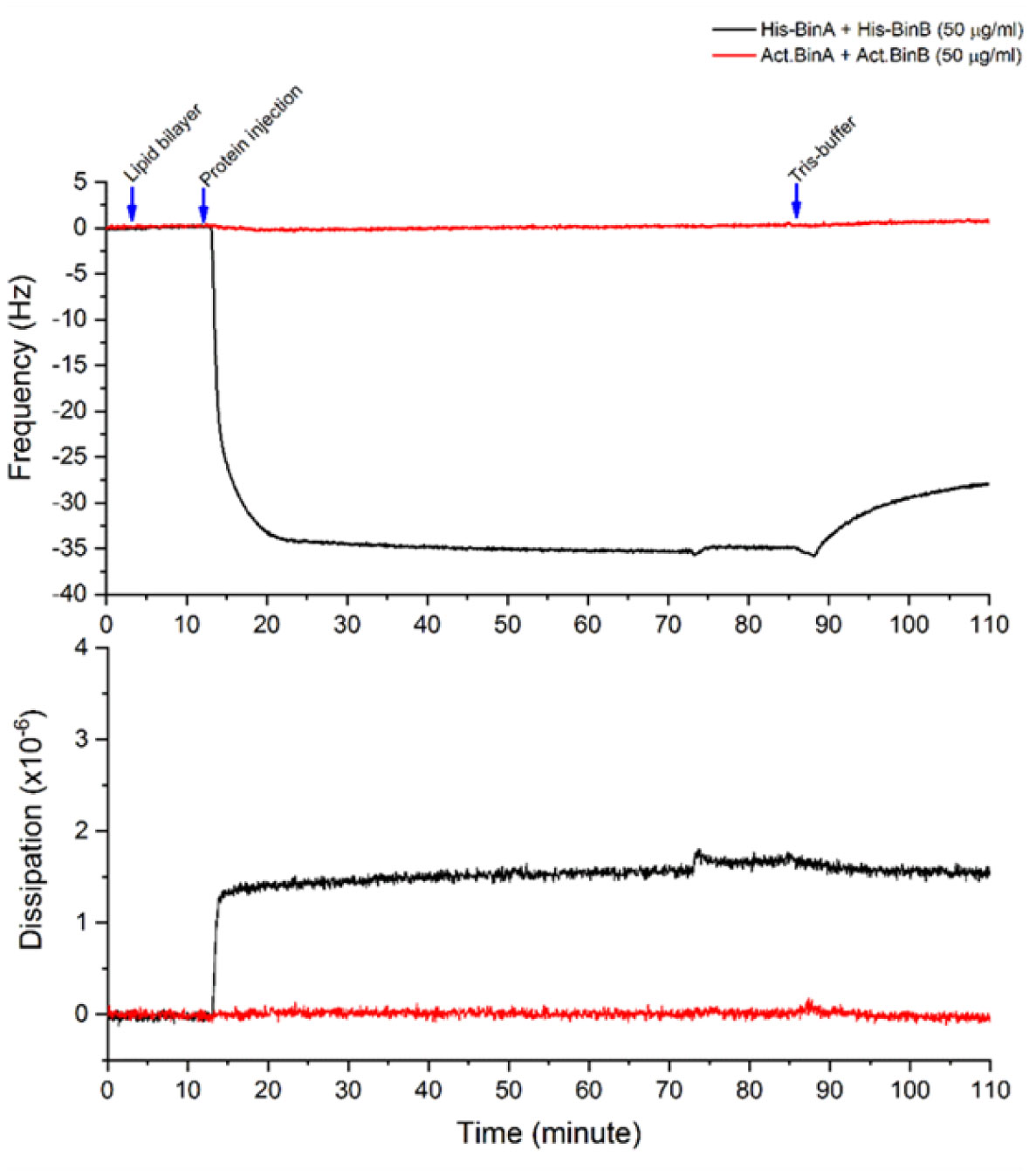
Binary toxin (Bin) is one of the bio-larvicidal toxin produced by Lysinibacillus sphaericus. Two component proteins, BinA and BinB toxins are required simultaneously to exert its larvicidal activity. The binary toxin has been proposed to act initially on the susceptible cell membrane. Here, the cell membrane binding of the binary toxin was imitated via specific histidine (His)-nickel ion (Ni2+) chelating. The N-terminal His-conjugated binary toxins (His-Bin) were attached onto the Ni2+-lipid bilayer surface besides its facilitating of purification process. The N-terminal conjugation of histidine tag did not interfere with the folding structure of both toxins. Subsequently, the attachment of binary toxins on the lipid membrane was successful with Ni2+-phosphatidylcholine (POPC)/phosphatidylethanolamine (POPE) bilayers (a model membrane that mimics the mosquito cell membrane) but not for Ni2+-phosphatidylcholine bilayer. However, His-BinA formed unstable attachment with Ni2+-POPC/POPE bilayers since it could be removed by buffer rinsing. In contrast, His-BinB required imidazole solution to detach from Ni2+-POPC/POPE. Particularly, His-BinB had higher binding affinity to Ni2+-ion than His-BinA. The lipid membrane attachment led to the initial finding that although BinA and BinB toxins share high homology structures, their capability for Ni2+ chelation was different. The local N-terminal structure of binary toxin seems to interfere the His-Ni2+ chelating of His-BinA.
Citation: Sudarat Tharad, Chontida Tangsongcharoen, Panadda Boonserm, José L. Toca-Herrera, Kanokporn Srisucharitpanit. Local conformations affect the histidine tag-Ni2+ binding affinity of BinA and BinB proteins[J]. AIMS Biophysics, 2020, 7(3): 133-143. doi: 10.3934/biophy.2020011
Binary toxin (Bin) is one of the bio-larvicidal toxin produced by Lysinibacillus sphaericus. Two component proteins, BinA and BinB toxins are required simultaneously to exert its larvicidal activity. The binary toxin has been proposed to act initially on the susceptible cell membrane. Here, the cell membrane binding of the binary toxin was imitated via specific histidine (His)-nickel ion (Ni2+) chelating. The N-terminal His-conjugated binary toxins (His-Bin) were attached onto the Ni2+-lipid bilayer surface besides its facilitating of purification process. The N-terminal conjugation of histidine tag did not interfere with the folding structure of both toxins. Subsequently, the attachment of binary toxins on the lipid membrane was successful with Ni2+-phosphatidylcholine (POPC)/phosphatidylethanolamine (POPE) bilayers (a model membrane that mimics the mosquito cell membrane) but not for Ni2+-phosphatidylcholine bilayer. However, His-BinA formed unstable attachment with Ni2+-POPC/POPE bilayers since it could be removed by buffer rinsing. In contrast, His-BinB required imidazole solution to detach from Ni2+-POPC/POPE. Particularly, His-BinB had higher binding affinity to Ni2+-ion than His-BinA. The lipid membrane attachment led to the initial finding that although BinA and BinB toxins share high homology structures, their capability for Ni2+ chelation was different. The local N-terminal structure of binary toxin seems to interfere the His-Ni2+ chelating of His-BinA.

| [1] |
Baumann L, Baumann P (1989) Expression in Bacillus subtilis of the 51- and 42-kilodalton mosquitocidal toxin genes of Bacillus sphaericus. Appl Environ Microbiol 55: 252-253. doi: 10.1128/AEM.55.1.252-253.1989

|
| [2] | Baumann P, Clark MA, Baumann L, et al. (1991) Bacillus sphaericus as a mosquito pathogen: properties of the organism and its toxins. Microbiol Mol Biol Rev 55: 425-436. |
| [3] |
Romão TP, de Melo Chalegre KD, Key S, et al. (2006) A second independent resistance mechanism to Bacillus sphaericus binary toxin targets its alpha-glucosidase receptor in Culex quinquefasciatus. FEBS J 273: 1556-1568. doi: 10.1111/j.1742-4658.2006.05177.x
|
| [4] |
Lacey LA, Undeen AH (1986) Microbial control of black flies and mosquitoes. Annu Rev Entomol 31: 265-296. doi: 10.1146/annurev.en.31.010186.001405
|
| [5] |
Oliveira CMF, Silva-Filha MH, Nielsen-Leroux C, et al. (2004) Inheritance and mechanism of resistance to Bacillus sphaericus in Culex quinquefasciatus (Diptera: Culicidae) from China and Brazil. J Med Entomol 41: 58-64. doi: 10.1603/0022-2585-41.1.58
|
| [6] |
Darboux I, Pauchet Y, Castella C, et al. (2002) Loss of the membrane anchor of the target receptor is a mechanism of bioinsecticide resistance. Proc Natl Acad Sci 99: 5830-5835. doi: 10.1073/pnas.092615399
|
| [7] |
Darboux I, Charles JF, Pauchet Y, et al. (2007) Transposon-mediated resistance to Bacillus sphaericus in a field-evolved population of Culex pipiens (Diptera: Culicidae). Cell Microbiol 9: 2022-2029. doi: 10.1111/j.1462-5822.2007.00934.x
|
| [8] |
Silva-Filha MH, Nielsen-LeRoux C, Charles JF (1999) Identification of the receptor for Bacillus sphaericus crystal toxin in the brush border membrane of the mosquito Culex pipiens (Diptera: Culicidae). Insect Biochem Mol Biol 29: 711-721. doi: 10.1016/S0965-1748(99)00047-8
|
| [9] |
Darboux I, Nielsen-LeRoux C, Charles JF, et al. (2001) The receptor of Bacillus sphaericus binary toxin in Culex pipiens (Diptera: Culicidae) midgut: molecular cloning and expression. Insect Biochem Mol Biol 31: 981-990. doi: 10.1016/S0965-1748(01)00046-7
|
| [10] |
Lekakarn H, Promdonkoy B, Boonserm P (2015) Interaction of Lysinibacillus sphaericus binary toxin with mosquito larval gut cells: Binding and internalization. J Invertebr Pathol 132: 125-131. doi: 10.1016/j.jip.2015.09.010
|
| [11] |
Tangsongcharoen C, Chomanee N, Promdonkoy B, et al. (2015) Lysinibacillus sphaericus binary toxin induces apoptosis in susceptible Culex quinquefasciatus larvae. J Invertebr Pathol 128: 57-63. doi: 10.1016/j.jip.2015.04.008
|
| [12] |
Tangsongcharoen C, Jupatanakul N, Promdonkoy B, et al. (2017) Molecular analysis of Culex quinquefasciatus larvae responses to Lysinibacillus sphaericus Bin toxin. PLoS One 12: e0175473. doi: 10.1371/journal.pone.0175473
|
| [13] |
Promdonkoy B, Promdonkoy P, Wongtawan B, et al. (2008) Cys31, Cys47, and Cys195 in BinA are essential for toxicity of a binary toxin from Bacillus sphaericus. Curr Microbiol 56: 334-338. doi: 10.1007/s00284-007-9065-9
|
| [14] |
Srisucharitpanit K, Yao M, Promdonkoy B, et al. (2014) Crystal structure of BinB: a receptor binding component of the binary toxin from Lysinibacillus sphaericus. Proteins: Structure, Function, and Bioinformatics 82: 2703-2712. doi: 10.1002/prot.24636
|
| [15] |
Colletier JP, Sawaya MR, Gingery M, et al. (2016) De novo phasing with X-ray laser reveals mosquito larvicide BinAB structure. Nature 539: 43-47. doi: 10.1038/nature19825
|
| [16] |
Srisucharitpanit K, Inchana P, Rungrod A, et al. (2012) Expression and purification of the active soluble form of Bacillus sphaericus binary toxin for structural analysis. Protein Expres Purif 82: 368-372. doi: 10.1016/j.pep.2012.02.009
|
| [17] |
Marheineke K, Grünewald S, Christie W, et al. (1998) Lipid composition of Spodoptera frugiperda (Sf9) and Trichoplusia ni (Tn) insect cells used for baculovirus infection. FEBS Lett 441: 49-52. doi: 10.1016/S0014-5793(98)01523-3
|
| [18] |
Knecht S, Ricklin D, Eberle AN, et al. (2009) Oligohis-tags: mechanisms of binding to Ni2+-NTA surfaces. J Mol Recognit 22: 270-279. doi: 10.1002/jmr.941
|
| [19] |
Chooduang S, Surya W, Torres J, et al. (2018) An aromatic cluster in Lysinibacillus sphaericus BinB involved in toxicity and proper in-membrane folding. Arch Biochem Biophys 660: 29-35. doi: 10.1016/j.abb.2018.10.006
|
Figures(4)

Sudarat Tharad, Chontida Tangsongcharoen, Panadda Boonserm, José L. Toca-Herrera, Kanokporn Srisucharitpanit. Local conformations affect the histidine tag-Ni2+ binding affinity of BinA and BinB proteins[J]. AIMS Biophysics, 2020, 7(3): 133-143. doi: 10.3934/biophy.2020011







 DownLoad:
DownLoad: