Citation: Constance J. Jeffery. Intracellular/surface moonlighting proteins that aid in the attachment of gut microbiota to the host[J]. AIMS Microbiology, 2019, 5(1): 77-86. doi: 10.3934/microbiol.2019.1.77

| [1] |
Jeffery CJ (1999) Moonlighting proteins. Trends Biochem Sci 24: 8–11. doi: 10.1016/S0968-0004(98)01335-8

|
| [2] | Chen C, Zabad S, Liu H, et al. (2018) MoonProt 2.0: an expansion and update of the moonlighting proteins database. Nucleic Acids Research 46: D640–D644. |
| [3] |
Kainulainen V, Korhonen TK (2014) Dancing to another tune-adhesive moonlighting proteins in bacteria. Biology 3: 178–204. doi: 10.3390/biology3010178
|
| [4] | Jeffery CJ (2018) Intracellular proteins moonlighting as bacterial adhesion factors. AIMS Microbiol 4: 362–376. |
| [5] |
Hennequin C, Porcheray F, Waligora-Dupriet A, et al. (2001) GroEL (Hsp60) of Clostridium difficile is involved in cell adherence. Microbiology 147: 87–96. doi: 10.1099/00221287-147-1-87
|
| [6] | Yamaguchi H, Osaki T, Kurihara N, et al. (1997) Heat-shock protein 60 homologue of Helicobacter pylori is associated with adhesion of H. pylori to human gastric epithelial cells. J Med Microbiol 46: 825–831. |
| [7] | Ensgraber M, Loos M (1992) A 66-kilodalton heat shock protein of Salmonella typhimurium is responsible for binding of the bacterium to intestinal mucus. Infect Immun 60: 3072–3078. |
| [8] | Hoffman PS, Garduno RA (1999) Surface-Associated heat shock proteins of Legionella pneumophila and Helicobacter pylori: Roles in pathogenesis and immunity. Infect Dis Obstet Gynecol 7: 58–63. |
| [9] |
Wampler JL, Kim KP, Jaradat Z, et al. (2004) Heat shock protein 60 acts as a receptor for the Listeria adhesion protein in Caco-2 cells. Infect Immun 72: 931–936. doi: 10.1128/IAI.72.2.931-936.2004
|
| [10] |
Jagadeesan B, Koo OK, Kim KP, et al. (2010) LAP, an alcohol acetaldehyde dehydrogenase enzyme in Listeria, promotes bacterial adhesion to enterocyte-like Caco-2 cells only in pathogenic species. Microbiology 156: 2782–2795. doi: 10.1099/mic.0.036509-0
|
| [11] |
Milohanic E, Pron B, Berche P, et al. (2000) Identification of new loci involved in adhesion of Listeria monocytogenes to eukaryotic cells. Microbiology 146: 731–739. doi: 10.1099/00221287-146-3-731
|
| [12] |
Castaldo C, Vastano V, Siciliano RA, et al. (2009) Surface displaced alfa-enolase of Lactobacillus plantarum is a fibronectin binding protein. Microb Cell Fact 8: 14. doi: 10.1186/1475-2859-8-14
|
| [13] |
Antikainen J, Kuparinen V, Lähteenmäki K, et al. (2007) pH-dependent association of enolase and glyceraldehyde-3-phosphate dehydrogenase of Lactobacillus crispatus with the cell wall and lipoteichoic acids. J Bacteriol 189: 4539–4543. doi: 10.1128/JB.00378-07
|
| [14] |
Patel DK, Shah KR, Pappachan A, et al. (2016) Cloning, expression and characterization of a mucin-binding GAPDH from Lactobacillus acidophilus. Int J Biol Macromol 91: 338–346. doi: 10.1016/j.ijbiomac.2016.04.041
|
| [15] |
Kainulainen V, Loimaranta V, Pekkala A, et al. (2012) Glutamine synthetase and glucose-6-phosphate isomerase are adhesive moonlighting proteins of Lactobacillus crispatus released by epithelial cathelicidin LL-37. J Bacteriol 194: 2509–2519. doi: 10.1128/JB.06704-11
|
| [16] |
Bergonzelli GE, Granato D, Pridmore RD, et al. (2006) GroEL of Lactobacillus johnsonii La1 (NCC 533) is cell surface associated: potential role in interactions with the host and the gastric pathogen Helicobacter pylori. Infect Immun 74: 425–434. doi: 10.1128/IAI.74.1.425-434.2006
|
| [17] |
Granato D, Bergonzelli GE, Pridmore RD, et al. (2004) Cell surface-associated elongation factor Tu mediates the attachment of Lactobacillus johnsonii NCC533 (La1) to human intestinal cells and mucins. Infect Immun 72:2160–2169. doi: 10.1128/IAI.72.4.2160-2169.2004
|
| [18] |
Kinoshita H, Uchida H, Kawai Y, et al. (2008) Cell surface Lactobacillus plantarum LA 318 glyceraldehyde-3-phosphate dehydrogenase (GAPDH) adheres to human colonic mucin. J Appl Microbiol 104: 1667–1674. doi: 10.1111/j.1365-2672.2007.03679.x
|
| [19] |
Candela M, Biagi E, Centanni M, et al. (2009) Bifidobacterial enolase, a cell surface receptor for human plasminogen involved in the interaction with the host. Microbiology 155: 3294–3303. doi: 10.1099/mic.0.028795-0
|
| [20] |
Candela M, Bergmann S, Vici M, et al. (2007) Binding of human plasminogen to Bifidobacterium. J Bacteriol 189: 5929–5936. doi: 10.1128/JB.00159-07
|
| [21] |
Crowe JD, Sievwright IK, Auld GC, et al. (2003) Candida albicans binds human plasminogen: identification of eight plasminogen-binding proteins. Mol Microbiol 47: 1637–1651. doi: 10.1046/j.1365-2958.2003.03390.x
|
| [22] | Gozalbo D, Gil-Navarro I, Azorin I, et al. (1998) The cell wall-associated glyceraldehyde-3-phosphate dehydrogenase of Candida albicans is also a fibronectin and laminin binding protein. Infect Immun 66: 2052–2059. |
| [23] |
Jong AY, Chen SH, Stins MF, et al. (2003) Binding of Candida albicans enolase to plasmin(ogen) results in enhanced invasion of human brain microvascular endothelial cells. J Med Microbiol 52: 615–622. doi: 10.1099/jmm.0.05060-0
|
| [24] |
Luo S, Hoffmann R, Skerka C, et al. (2013) Glycerol-3-phosphate dehydrogenase 2 is a novel factor H-, factor H-like protein 1-,and plasminogen binding surface protein of Candida albicans. J Infect Dis 207: 594–603. doi: 10.1093/infdis/jis718
|
| [25] |
Lesiak-Markowicz I, Vogl G, Schwarzmuller T, et al. (2011) Candida albicans Hgt1p, a multifunctional evasion molecule: complement inhibitor, CR3 analogue,and human immunodeficiency virus-binding molecule. J Infect Dis 204: 802–809. doi: 10.1093/infdis/jir455
|
| [26] |
Yang W, Li E, Kairong T, et al. (1994) Entamoeba histolytica has an alcohol dehydrogenase homologous to themultifunctional adhE gene product of Escherichia coli. Mol Biochem Parasitol 64: 253–60. doi: 10.1016/0166-6851(93)00020-A
|
| [27] |
Jin S, Song YC, Emili A, et al. (2003) JlpA of Campylobacter jejuni interacts with surface-exposed heat shock protein 90-alpha and triggers signaling pathways leading to the activation of NF-kappaB and p38 MAP kinase in epithelial cells. Cell Microbiol 5: 165–74. doi: 10.1046/j.1462-5822.2003.00265.x
|
| [28] |
Raymond BB, Djordjevic S (2015) Exploitation of plasmin(ogen) by bacterial pathogens of veterinary significance. Vet Microbiol 178: 1–13. doi: 10.1016/j.vetmic.2015.04.008
|
| [29] | Collen D, Verstraete M (1975) Molecular biology of human plasminogen II Metabolism in physiological and some pathological conditions in man. Thromb Diath Haemorrh 34: 403–408. |
| [30] |
Dano K, Andreasen PA, Grondahl-Hansen J, et al. (1985) Plasminogen activators, tissue degradation, and cancer. Adv Cancer Res 44: 139–266. doi: 10.1016/S0065-230X(08)60028-7
|
| [31] |
Wang W, Jeffery CJ (2016) An analysis of surface proteomics results reveals novel candidates for intracellular/surface moonlighting proteins in bacteria. Mol Biosyst 12: 1420–1431. doi: 10.1039/C5MB00550G
|
| [32] |
Han MJ, Lee SY, Hong SH (2012) Comparative analysis of envelope proteomes in Escherichia coli B and K-12 strains. J Microbiol Biotechnol 22: 470–478. doi: 10.4014/jmb.1110.10080
|
| [33] |
Bøhle LA, Riaz T, Egge-Jacobsen W, et al. (2011) Identification of surface proteins in Enterococcus faecalis V583. BMC Genomics 12: 135. doi: 10.1186/1471-2164-12-135
|
| [34] |
Garcia-del Portillo F, Calvo E, D'Orazio V, et al. (2011) Association of ActA to peptidoglycan revealed by cell wall prteomics of intracellular Listeria monocytogenes. J Biol Chem 286: 34675–34689. doi: 10.1074/jbc.M111.230441
|
| [35] |
Amblee V, Jeffery CJ (2015) Physical features of intracellular proteins that moonlight on the cell surface. PLoS One 10: e0130575. doi: 10.1371/journal.pone.0130575
|
| [36] |
Matsuoka K, Kanai T (2015) The gut microbiota and inflammatory bowel disease. Semin Immunopathol 37: 47–55. doi: 10.1007/s00281-014-0454-4
|
| [37] | Dahlhamer JM, Zammitti EP, Ward BW, et al. (2016) Prevalence of inflammatory bowel disease among adults aged ≥18 years-United States. MMWR 65:1166–1169. |
| [38] |
Kinoshita H, Ohuchi S, Arakawa K, et al. (2016) Isolation of lactic acid bacteria bound to the porcine intestinal mucosa and an analysis of their moonlighting adhesins. Biosci Microbiota Food Health 35:185–196. doi: 10.12938/bmfh.16-012
|
| [39] |
Celebioglu HU, Olesen SV, Prehn K, et al. (2017) Mucin-and carbohydrate-stimulated adhesion and subproteome changes of the probiotic bacterium Lactobacillus acidophilus NCFM. J Proteomics. 163: 102–110. doi: 10.1016/j.jprot.2017.05.015
|
| [40] | Zhu D, Sun Y, Liu F, et al. (2016) Identification of surface-associated proteins of Bifidobacterium animalis ssp. Lactis KLDS 2.0603 by enzymatic shaving. J Dairy Sci. 99: 5155–5172. |
| [41] | Celebioglu HU, Svensson B (2017) Exo-and surface proteomes of the probiotic bacterium Lactobacillus acidophilus NCFM. Proteomics. 17: 11. |
| [42] |
Celebioglu HU, Delsoglio M, Brix S, et al. (2018) Plant polyphenols stimulate adhesion to intestinal mucosa and induce proteome changes in the probiotic Lactobacillus acidophilus NCFM. Mol Nutr Food Res. 62: 1700638. doi: 10.1002/mnfr.201700638
|
| [43] |
Celebioglu HU, Ejby M, Majumder A, et al. (2016) Differential proteome and cellular adhesion analyses of the probiotic bacterium Lactobacillus acidophilus NCFM grown on raffinose - an emerging prebiotic. Proteomics. 16: 1361–1375. doi: 10.1002/pmic.201500212
|
| [44] |
Pérez Montoro B, Benomar N, Caballero Gómez N, et al. (2018) Proteomic analysis of Lactobacillus pentosus for the identification of potential markers of adhesion and other probiotic features. Food Res Int. 111: 58–66. doi: 10.1016/j.foodres.2018.04.072
|
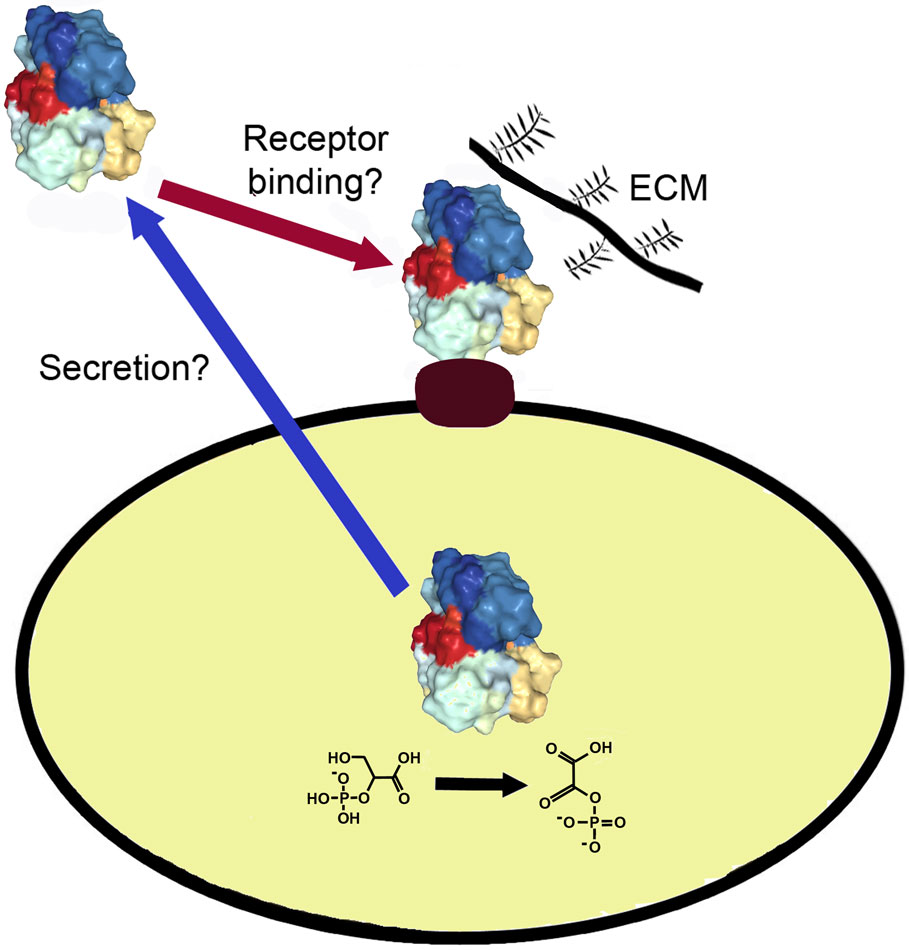
Figures(1) / Tables(1)

Constance J. Jeffery. Intracellular/surface moonlighting proteins that aid in the attachment of gut microbiota to the host[J]. AIMS Microbiology, 2019, 5(1): 77-86. doi: 10.3934/microbiol.2019.1.77







 DownLoad:
DownLoad: