Citation: Hayet Saidani, Daria Grobys, Marc Léonetti, Hanna Kmita, Fabrice Homblé. Towards understanding of plant mitochondrial VDAC proteins: an overview of bean (Phaseolus) VDAC proteins[J]. AIMS Biophysics, 2017, 4(1): 43-62. doi: 10.3934/biophy.2017.1.43

| [1] |
Delgado-Salinas A, Bibler R, Lavin M (2006) Phylogeny of the genus phaseolus (leguminosae): a recent diversification in an ancient landscape. Syst Bot 31: 779–791. doi: 10.1600/036364406779695960

|
| [2] |
Chacón S MI, Pickersgill B, Debouck DG, et al. (2007) Phylogeographic analysis of the chloroplast DNA variation in wild common bean (Phaseolus vulgaris L.) in the Americas. Plant Syst Evol 266: 175–195. doi: 10.1007/s00606-007-0536-z
|
| [3] |
Parreira JR, Bouraada J, Fitzpatrick MA, et al. (2016) Differential proteomics reveals the hallmarks of seed development in common bean (Phaseolus vulgaris L.). J Proteomics 143: 188–198. doi: 10.1016/j.jprot.2016.03.002
|
| [4] |
Woodstock LW, Pollock BM (1965) Physiological predetermination: imbibition, respiration, and growth of lima bean seeds. Science 150: 1031–1032. doi: 10.1126/science.150.3699.1031
|
| [5] | Palmieri F, Agrimi G, Blanco E, et al. (2006) Identification of mitochondrial carriers in Saccharomyces cerevisiae by transport assay of reconstituted recombinant proteins. Biochim Biophys Acta 1757: 1249–1262. |
| [6] |
Inoue I, Nagase H, Kishi K, et al. (1991) ATP-sensitive K+ channel in the mitochondrial inner membrane. Nature 352: 244–247. doi: 10.1038/352244a0
|
| [7] |
Siemen D, Loupatatzis C, Borecky J, et al. (1999) Ca2+-activated K channel of the BK-Type in the inner mitochondrial membrane of a human glioma cell line. Biochem Biophys Res Commun 257: 549–554. doi: 10.1006/bbrc.1999.0496
|
| [8] |
Szabò I, Bock J, Jekle A, et al. (2005) A novel potassium channel in lymphocyte mitochondria. J Biol Chem 280: 12790–12798. doi: 10.1074/jbc.M413548200
|
| [9] |
Pang K, Li Y, Liu M, et al. (2013) Inventory and general analysis of the ATP-binding cassette (ABC) gene superfamily in maize (Zea mays L.). Gene 526: 411–428. doi: 10.1016/j.gene.2013.05.051
|
| [10] |
Srinivasan V, Pierik AJ, Lill R (2014) Crystal structures of nucleotide-free and glutathione-bound mitochondrial ABC transporter atm1. Science 343: 1137–1140. doi: 10.1126/science.1246729
|
| [11] |
Homblé F, Krammer E-M, Prévost M (2012) Plant VDAC: Facts and speculations. Biochim Biophys Acta-Biomembr 1818: 1486–1501. doi: 10.1016/j.bbamem.2011.11.028
|
| [12] |
Marmagne A, Rouet M-A, Ferro M, et al. (2004) Identification of new intrinsic proteins in Arabidopsis plasma membrane proteome. Mol Cell Proteomics 3: 675–691. doi: 10.1074/mcp.M400001-MCP200
|
| [13] |
Robert N, D’Erfurth I, Marmagne A, et al. (2012) Voltage-dependent-anion-channels (VDACs) in Arabidopsis have a dual localization in the cell but show a distinct role in mitochondria. Plant Mol Biol 78: 431–446. doi: 10.1007/s11103-012-9874-5
|
| [14] |
De Pinto V, Messina A, Lane DJR, et al. (2010) Voltage-dependent anion-selective channel (VDAC) in the plasma membrane. FEBS Lett 584: 1793–1799. doi: 10.1016/j.febslet.2010.02.049
|
| [15] |
Shoshan-Barmatz V, Pinto V De, Zweckstetter M, et al. (2010) VDAC, a multi-functional mitochondrial protein regulating cell life and death. Mol Aspects Med 31: 227–285. doi: 10.1016/j.mam.2010.03.002
|
| [16] | Baker MA, Lane DJR, Ly JD, et al. (2004) VDAC1 is a transplasma membrane NADH-ferricyanide reductase. J Biol Chem 279: 4811–4819. |
| [17] |
Parsons DF, Bonner J, Verboon JG (1965) Electron microscopy of isolated plant mitochondria and plastids using both the thin-section and the negative-staining techniques. Can J Bot 43: 647–655. doi: 10.1139/b65-072
|
| [18] |
Mannella CA, Bonner WD (1975) X-ray diffraction from oriented outer mitochondrial membranes. Biochim Biophys Acta-Biomembr 413: 226–233. doi: 10.1016/0005-2736(75)90106-6
|
| [19] | Zalman LS, Nikaido H, Kagawa Y (1980) Mitochondrial outer membrane contains a protein producing nonspecific diffusion channels. J Biol Chem 255: 1771–1774. |
| [20] |
Schmutz J, McClean PE, Mamidi S, et al. (2014) A reference genome for common bean and genome-wide analysis of dual domestications. Nat Genet 46: 707–713. doi: 10.1038/ng.3008
|
| [21] |
Abrecht H, Wattiez R, Ruysschaert JM, et al. (2000) Purification and characterization of two voltage-dependent anion channel isoforms from plant seeds. Plant Physiol 124: 1181–1190. doi: 10.1104/pp.124.3.1181
|
| [22] |
Abrecht H, Goormaghtigh E, Ruysschaert JM, et al. (2000) Structure and orientation of two voltage-dependent anion-selective channel isoforms—An attenuated total reflection Fourier-transform infrared spectroscopy study. J Biol Chem 275: 40992–40999. doi: 10.1074/jbc.M006437200
|
| [23] | Homblé F, Mlayeh L, Léonetti M (2010) Planar lipid bilayers for electrophysiology of membrane-active peptides, In: Membr Pept Methods Results Struct Funct Electrophysiol, IUL: La Jolla, 273–307. |
| [24] |
Mlayeh L, Chatkaew S, Léonetti M, et al. (2010) Modulation of plant mitochondrial VDAC by phytosterols. Biophys J 99: 2097–2106. doi: 10.1016/j.bpj.2010.07.067
|
| [25] | Schein SJ, Colombini M, Finkelstein A (1976) Reconstitution in planar lipid bilayers of a voltage-dependent anion-selective channel obtained from paramecium mitochondria. J Membr Biol 30: 99–120. |
| [26] |
Rostovtseva TK, Colombini M (1996) ATP flux is controlled by a voltage-gated channel from the mitochondrial outer membrane. J Biol Chem 271: 28006–28008. doi: 10.1074/jbc.271.45.28006
|
| [27] |
Schmid A, Kromer S, Heldt HW, et al. (1992) Identification of two general diffusion channels in the outer membrane of pea mitochondria. Biochim Biophys Acta 1112: 174–180. doi: 10.1016/0005-2736(92)90389-4
|
| [28] | Heins L, Mentzel H, Schmid A, et al. (1994) Biochemical, molecular, and functional characterization of porin isoforms from potato mitochondria. J Biol Chem 269: 26402–26410. |
| [29] |
Blumenthal A, Kahn K, Beja O, et al. (1993) Purification and characterization of the voltage-dependent anion-selective channel protein from wheat mitochondrial membranes. Plant Physiol 101: 579–587. doi: 10.1104/pp.101.2.579
|
| [30] | Smack DP, Colombini M (1985) Voltage-dependent channels found in the membrane fraction of corn mitochondria. Plant Physiol 79: 1094–1097. |
| [31] |
Aljamal JA, Genchi G, De Pinto V, et al. (1993) Purification and characterization of porin from corn (Zea mays L.) mitochondria. Plant Physiol 102: 615–621. doi: 10.1104/pp.102.2.615
|
| [32] |
Wunder UR, Colombini M (1991) Patch clamping VDAC in liposomes containing whole mitochondrial membranes. J Membr Biol 123: 83–91. doi: 10.1007/BF01993966
|
| [33] |
Levadny V, Colombini M, Li XX, et al. (2002) Electrostatics explains the shift in VDAC gating with salt activity gradient. Biophys J 82: 1773–1783. doi: 10.1016/S0006-3495(02)75528-8
|
| [34] |
Bayrhuber M, Meins T, Habeck M, et al. (2008) Structure of the human voltage-dependent anion channel. Proc Natl Acad Sci USA 105: 15370–15375. doi: 10.1073/pnas.0808115105
|
| [35] |
Hiller S, Garces RG, Malia TJ, et al. (2008) Solution structure of the integral human membrane protein VDAC-1 in detergent micelles. Science 321: 1206–1210. doi: 10.1126/science.1161302
|
| [36] | Ujwal R, Cascio D, Colletier JP, et al. (2008) The crystal structure of mouse VDAC1 at 2.3 A resolution reveals mechanistic insights into metabolite gating. Proc Natl Acad Sci USA 105: 17742–17747. |
| [37] |
Schredelseker J, Paz A, Lopez CJ, et al. (2014) High resolution structure and double electron-electron resonance of the zebrafish voltage-dependent anion channel 2 reveal an oligomeric population. J Biol Chem 289: 12566–12577. doi: 10.1074/jbc.M113.497438
|
| [38] |
Schneider R, Etzkorn M, Giller K, et al. (2010) The native conformation of the human VDAC1 N terminus. Angew Chemie Int Ed 49: 1882–1885. doi: 10.1002/anie.200906241
|
| [39] |
Guardiani C, Scorciapino MA, Amodeo GF, et al. (2015) The N-terminal peptides of the three human isoforms of the mitochondrial voltage-dependent anion channel have different helical propensities. Biochemistry 54: 5646–5656. doi: 10.1021/acs.biochem.5b00469
|
| [40] |
De Pinto V, Tomasello F, Messina A, et al. (2007) Determination of the conformation of the human VDAC1 N-terminal peptide, a protein moiety essential for the functional properties of the pore. Chem Bio Chem 8: 744–756. doi: 10.1002/cbic.200700009
|
| [41] |
Colombini M (2009) The published 3D structure of the VDAC channel: native or not? Trends Biochem Sci 34: 382–389. doi: 10.1016/j.tibs.2009.05.001
|
| [42] | Colombini M (2012) VDAC structure, selectivity, and dynamics. Biochim Biophys Acta-Biomembr 1818: 1457–1465. |
| [43] |
Lee K Il, Rui H, Pastor RW, et al. (2011) Brownian dynamics simulations of ion transport through the VDAC. Biophys J 100: 611–619. doi: 10.1016/j.bpj.2010.12.3708
|
| [44] |
Choudhary OP, Ujwal R, Kowallis W, et al. (2010) The electrostatics of VDAC: implications for selectivity and gating. J Mol Biol 396: 580–592. doi: 10.1016/j.jmb.2009.12.006
|
| [45] |
Choudhary OP, Paz A, Adelman JL, et al. (2014) Structure-guided simulations illuminate the mechanism of ATP transport through VDAC1. Nat Struct Mol Biol 21: 626–632. doi: 10.1038/nsmb.2841
|
| [46] |
Hiller S, Abramson J, Mannella C, et al. (2010) The 3D structures of VDAC represent a native conformation. Trends Biochem Sci 35: 514–521. doi: 10.1016/j.tibs.2010.03.005
|
| [47] |
Noskov SY, Rostovtseva TK, Bezrukov SM (2013) ATP Transport through VDAC and the VDAC-tubulin complex probed by equilibrium and nonequilibrium MD simulations. Biochemistry 52: 9246–9256. doi: 10.1021/bi4011495
|
| [48] | Rui H, Lee K Il, Pastor RW, et al. (2011) Molecular dynamics studies of ion permeation in VDAC. Biophys J 100: 602–610. |
| [49] |
Young MJ, Bay DC, Hausner G, et al. (2007) The evolutionary history of mitochondrial porins. BMC Evol Biol 7: 31. doi: 10.1186/1471-2148-7-31
|
| [50] | Kutik S, Stojanovski D, Becker L, et al. (2008) Dissecting membrane insertion of mitochondrial beta-barrel proteins. Cell 132: 1011–1024. |
| [51] |
Imai K, Fujita N, Gromiha MM, et al. (2011) Eukaryote-wide sequence analysis of mitochondrial beta-barrel outer membrane proteins. BMC Genomics 12: 79. doi: 10.1186/1471-2164-12-79
|
| [52] | Jores T, Klinger A, Grosz LE, et al. (2016) Characterization of the targeting signal in mitochondrial [beta]-barrel proteins. Nat Commun 7. |
| [53] |
Shanmugavadivu B, Apell HJ, Meins T, et al. (2007) Correct folding of the beta-barrel of the human membrane protein VDAC requires a lipid bilayer. J Mol Biol 368: 66–78. doi: 10.1016/j.jmb.2007.01.066
|
| [54] |
Shi Y, Jiang C, Chen Q, et al. (2003) One-step on-column affinity refolding purification and functional analysis of recombinant human VDAC1. Biochem Biophys Res Commun 303: 475–482. doi: 10.1016/S0006-291X(03)00359-0
|
| [55] | Koppel DA, Kinnally KW, Masters P, et al. (1998) Bacterial expression and characterization of the mitochondrial outer membrane channel. Effects of n-terminal modifications. J Biol Chem 273: 13794–13800. |
| [56] |
Shao L, Kinnally KW, Mannella CA (1996) Circular dichroism studies of the mitochondrial channel, VDAC, from Neurospora crassa. Biophys J 71: 778–786. doi: 10.1016/S0006-3495(96)79277-9
|
| [57] | Smeyers M, Léonetti M, Goormaghtigh E, et al. (2003) Structure and function of plant membrane ion channels reconstituted in planar lipid bilayers, In: Membr Sci Technol—Planar lipid bilayers their appl, Elsevier, 449–478. |
| [58] |
Villinger S, Briones R, Giller K, et al. (2010) Functional dynamics in the voltage-dependent anion channel. Proc Natl Acad Sci 107: 22546–22551. doi: 10.1073/pnas.1012310108
|
| [59] |
Blachly-Dyson E, Peng S, Colombini M, et al. (1990) Selectivity changes in site-directed mutants of the VDAC ion channel: structural implications. Science 247: 1233–1236. doi: 10.1126/science.1690454
|
| [60] |
Zambrowicz EB, Colombini M (1993) Zero-current potentials in a large membrane channel: a simple theory accounts for complex behavior. Biophys J 65: 1093–1100. doi: 10.1016/S0006-3495(93)81148-2
|
| [61] | Krammer E-M, Saidani H, Prévost M, et al. (2014) Origin of ion selectivity in Phaseolus coccineus VDAC mitochondrial channel. Mitochondrion 19B: 206–213. |
| [62] |
Rosenquist M, Sehnke P, Ferl RJ, et al. (2000) Evolution of the 14-3-3 protein family: does the large number of isoforms in multicellular organisms reflect functional specificity? J Mol Evol 51: 446–458. doi: 10.1007/s002390010107
|
| [63] | Gasteiger E, Hoogland C, Gattiker A, et al. (2005) Protein identification and analysis tools on the ExPASy server, In: Walker JM, Proteomics Protoc, Totowa: Humana Press, 571–607 |
| [64] |
Schleiff E, Eichacker LA, Eckart K, et al. (2003) Prediction of the plant beta-barrel proteome: A case study of the chloroplast outer envelope. Protein Sci 12: 748–759. doi: 10.1110/ps.0237503
|
| [65] | Kumar S, Stecher G, Tamura K (2016) MEGA7: molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol Biol Evol 33. |
| [66] |
Smith MD, Petrak M, Boucher PD, et al. (1995) Lysine residues at positions 234 and 236 in yeast porin are involved in its assembly into the mitochondrial outer membrane. J Biol Chem 270: 28331–28336. doi: 10.1074/jbc.270.47.28331
|
| [67] |
Tateda C, Watanabe K, Kusano T, et al. (2011) Molecular and genetic characterization of the gene family encoding the voltage-dependent anion channel in Arabidopsis. J Exp Bot 62: 4773–4785. doi: 10.1093/jxb/err113
|
| [68] | Edgar RC (2004) MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res 32: 1792–1797. |
| [69] |
Schneider TD, Stephens RM (1990) Sequence logos: a new way to display consensus sequences. Nucleic Acids Res 18: 6097–6100. doi: 10.1093/nar/18.20.6097
|
| [70] |
Crooks GE, Hon G, Chandonia J-M, et al. (2004) WebLogo: a sequence logo generator. Genome Res 14: 1188–1190. doi: 10.1101/gr.849004
|
| [71] |
Biasini M, Bienert S, Waterhouse A, et al. (2014) SWISS-MODEL: modelling protein tertiary and quaternary structure using evolutionary information. Nucleic Acids Res 42: W252–W258. doi: 10.1093/nar/gku340
|
| [72] |
Yang Y, Faraggi E, Zhao H, et al. (2011) Improving protein fold recognition and template-based modeling by employing probabilistic-based matching between predicted one-dimensional structural properties of query and corresponding native properties of templates. Bioinformatics 27: 2076–2082. doi: 10.1093/bioinformatics/btr350
|
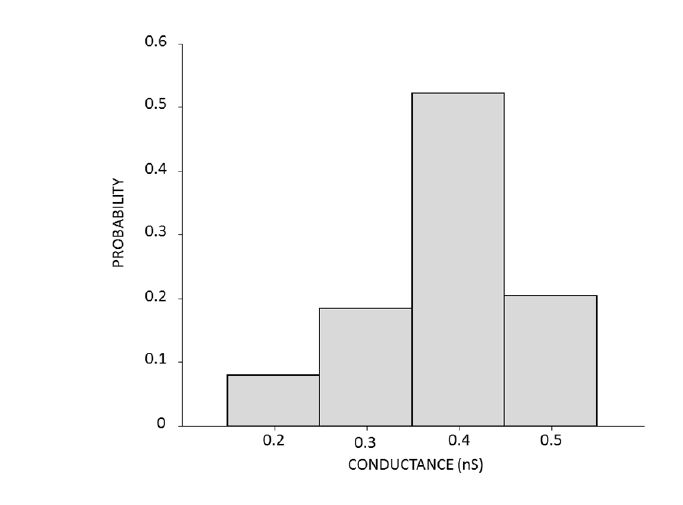
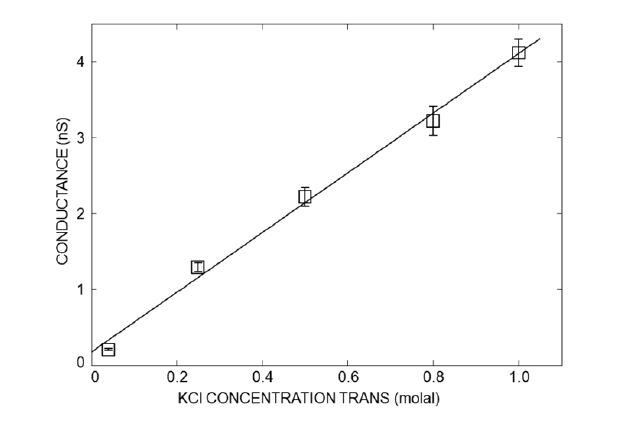
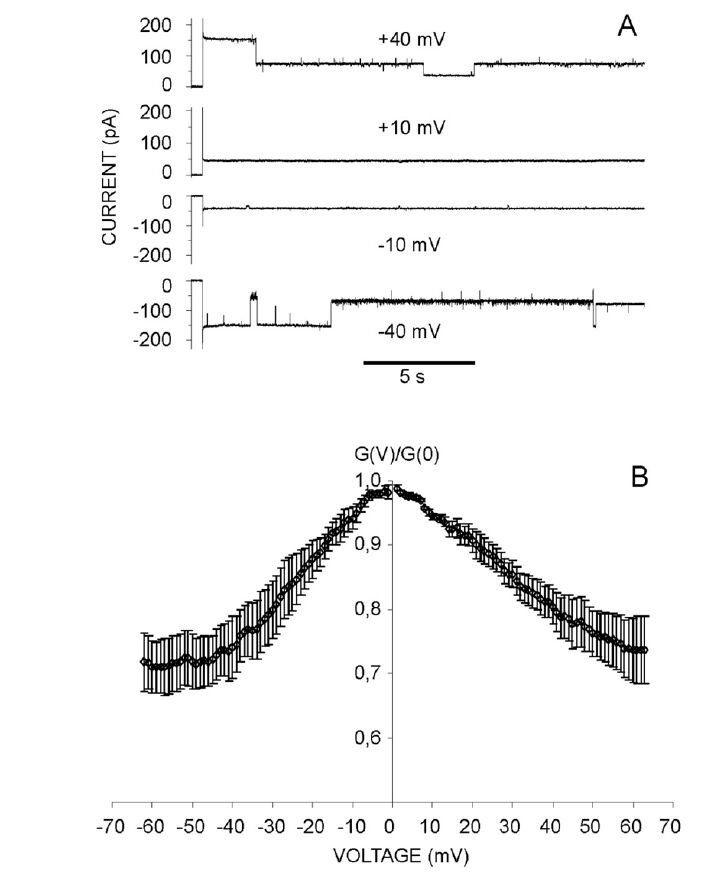
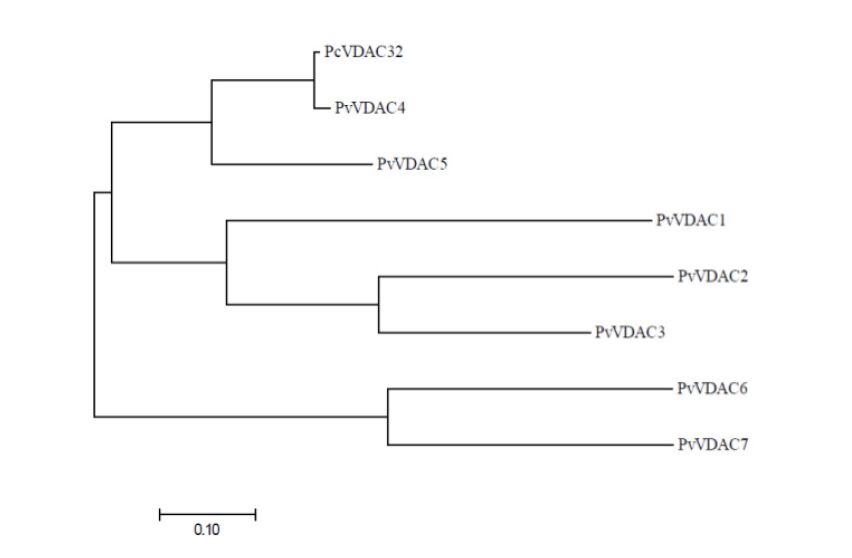


Figures(10) / Tables(3)

Hayet Saidani, Daria Grobys, Marc Léonetti, Hanna Kmita, Fabrice Homblé. Towards understanding of plant mitochondrial VDAC proteins: an overview of bean (Phaseolus) VDAC proteins[J]. AIMS Biophysics, 2017, 4(1): 43-62. doi: 10.3934/biophy.2017.1.43







 DownLoad:
DownLoad: