Citation: Tuan Syaripah Najihah, Mohd Hafiz Ibrahim, A'fifah Abd Razak, Rosimah Nulit, Puteri Edaroyati Megat Wahab. Effects of water stress on the growth, physiology and biochemical properties of oil palm seedlings[J]. AIMS Agriculture and Food, 2019, 4(4): 854-868. doi: 10.3934/agrfood.2019.4.854

| [1] | Kushairi A, Singh R, Ong-Abdullah M (2017) The oil palm industry in Malaysia: Thriving with transformative technologies. J Oil Palm Res 29: 431-439. |
| [2] | MPOB (2019) Monthly export of oil palm Products-2018. Available from: http://bepi.mpob.gov.my/index.php/en/statistics/export/192-export-2018/869 monthly- export-of-oil-palm-products-2018.html. |
| [3] |
Paterson RRM, Lima N (2018) Climate change affecting oil palm agronomy, and oil palm cultivation increasing climate change, require amelioration. Ecol Evol 8: 452-461. doi: 10.1002/ece3.3610

|
| [4] | Paterson RRM, Kumar L, Taylor S, et al. (2015) Future climate effects on suitability for growth of oil palms in Malaysia and Indonesia. Sci Rep 5: 14457. |
| [5] | Siwar C, Ahmed F, Begum RA (2013) Climate change, agriculture and food security issues: Malaysian perspective. J Food, Agric Environ 11: 1118-1123. |
| [6] | Basiron Y (2007) Palm oil production through sustainable plantations. Eur J Lipid Sci Technol 109: 289-295. |
| [7] | Oettli P, Behera SK, Yamagata T (2018) Climate-based predictability of oil palm tree yield in Malaysia. Sci Rep 8: 2271. |
| [8] | Intergovernmental Panel on Climate Change (IPCC) (2014) Climate change 2014: Synthesis report: Contribution of Working Groups I, Ⅱ and Ⅲ to the fifth assessment report of the Intergovernmental Panel on Climate Change. Available from: http://www.ipcc.ch/report/ar5/syr/. |
| [9] | Sheffield J, Wood EF, Roderick ML (2012) Little change in global drought over the past 60 years. Nature 491: 435-438. |
| [10] | Kamil NN, Omar SF (2017) The impact of El Niño and La Niña on Malaysian palm oil industry. Oil Palm Bull 74: 1-6. |
| [11] |
Shidiq IPA, Ismail MH, Kamarudin N, et al. (2018) Projections on future impact and vulnerability of climate change towards rubber areas in Peninsular Projections on future impact and vulnerability of climate change towards rubber areas in Peninsular Malaysia. IOP Conference Series: Earth and Environmental Science 169: 012053. doi: 10.1088/1755-1315/169/1/012053
|
| [12] | Amirul HA, Tui L, Yaw S, et al. (2016) Impact of El Niño on palm oil production. Planter 92: 789-806. |
| [13] | Adnan H (2015) El Nino dry weather to lead to rise in the palm oil price. Available from: https://www.thestar.com.my/business/business-news/2015/09/28/the-return-of-el-nino/. |
| [14] | Culman M, De Farias CM, Bayona C, et al. (2019) Using agrometeorological data to assist irrigation management in oil palm crops: A decision support method and results from crop model simulation. Agric Water Manage 213: 1047-1062. |
| [15] | Noor MRM, Harun MH (2004) Importance of water use efficiency (WUE) in oil palm productivity. Oil Palm Bull 48: 24-30. |
| [16] | Silva PA, Oliveira IV, Rodrigues KCB, et al. (2016) Leaf gas exchange and multiple enzymatic and non-enzymatic antioxidant strategies related to drought tolerance in two oil palm hybrids. Trees-Struct Funct 30: 203-214. |
| [17] | Putra ETS, Issukindarsyah, Taryono, et al. (2015) Physiological responses of oil palm seedlings to the drought stress using boron and silicon applications. J Agron 14: 49-61. |
| [18] | Tan CC (2011) Nursery practices for production of superior oil palm planting materials. Agronomic Princ Pract Oil Palm Cultiv 145-169. |
| [19] | Campos H, Trejo C, Peña-Valdivia CB, et al. (2014) Stomatal and non-stomatal limitations of bell pepper (Capsicum annuum L.) plants under water stress and re-watering: Delayed restoration of photosynthesis during recovery. Environ Exp Bot 98: 56-64. |
| [20] | Hopkins WG, Huner NPA (1999) Introduction to plant physiology. John Wiley and Sons, New York, 25-50. |
| [21] | Liu F, Jensen CR, Shahanzari A, et al. (2005) ABA-regulated stomatal control and photosynthetic water use efficiency of potato (Solanum tuberosum L.) during progressive soil drying. Plant Sci 168: 831-836. |
| [22] | Cha-um S, Yamada N, Takabe T, et al. (2013) Physiological features and growth characters of oil palm (Elaeis guineensis Jacq.) in response to reduced water-deficit and rewatering. Aust J Crop Sci 7: 432-439. |
| [23] | Suresh K, Nagamani C, Ramachandrudu K, et al. (2010) Gas-exchange characteristics, leaf water potential and chlorophyll a fluorescence in oil palm (Elaeis guineensis Jacq.) seedlings under water stress and recovery. Photosynthetica 48: 430-436. |
| [24] | Noor MRM, Harun MH, Jantan NM (2011) Physiological plant stress and responses in oil palm. Oil Palm Bull 62: 25-32. |
| [25] | Klapwijk D, Lint PJAL (1974) Fresh weight and flowering of tomato plants as influenced by container type and watering conditions. Acta Hortic 23: 237-248. |
| [26] | MPOB (2017) Oil palm Vegetative Measurements Manual. Kuala Lumpur. Malaysian Palm Oil Board, 50-65. |
| [27] |
Weatherly P (1950) Studies in the water relations of the cotton plant. New Phytol 49: 81-97. doi: 10.1111/j.1469-8137.1950.tb05146.x
|
| [28] | Zainuren NF, Ibrahim MH, Abdullah CAC, et al. (2019) Growth, carbon assimilation and quality of kesum (Persicaria minor) as exposed to zinc oxide nanoparticles. Annu Res. Rev Biol 31: 1-15. |
| [29] |
Jaafar HZE, Ibrahim MH, Fakri NFM (2012) Impact of soil field water capacity on secondary metabolites, phenylalanine ammonia-lyase (PAL), malondialdehyde (MDA) and photosynthetic responses of Malaysian Kacip Fatimah (Labisia pumila Benth). Molecules 17: 7305-7322. doi: 10.3390/molecules17067305
|
| [30] | Ibrahim MH, Jaafar HZE, Rahmat A, et al. (2012) Involvement of nitrogen on flavonoids, glutathione, anthocyanin, ascorbic acid and antioxidant activities of Malaysian medicinal plant Labisia pumila Blume (Kacip Fatimah). Int J Mol Sci 13: 393-408. |
| [31] | Bates LS, Waldren RP, Teare ID (1973) Rapid determination of free proline for water-stress studies. Plant Soil 39: 205-207. |
| [32] | Abdul Jaleel C, Manivannan P, Wahid A, et al. (2009) Drought stress in plants: A Review on morphological characteristics and pigments composition. Int J Agric Biol 11: 100-105. |
| [33] | Farooq M, Hussain M, Wahid A, et al. (2012) Drought stress in plants: An overview BT-Plant responses to drought stress: From morphological to molecular features. In: Aroca R, Springer Berlin Heidelberg, Berlin, Heidelberg, 1-33. |
| [34] | Anjum S, Xie X, Wang L, et al. (2011) Morphological, physiological and biochemical responses of plants to drought stress. Afr J Agric Res 6: 2026-2032. |
| [35] | Rivera-Mendes YD, Cuenca JC, Romero HM (2016) Physiological responses of oil palm (Elaeis guineensis Jacq.) seedlings under different water soil conditions. Agronomía Colombiana 34: 163-171. |
| [36] | Boutraa T, Akhkha A, Al-Shoaibi AA, et al. (2010) Effect of water stress on growth and water use efficiency (WUE) of some wheat cultivars (Triticum durum) grown in Saudi Arabia. J Taibah Univ Sci 3: 39-48. |
| [37] | Chartzoulakis K, Noitsakis B, Therios I (1993) Photosynthesis, plant growth and dry matter distribution in kiwifruit as influenced by water deficits. Irrig Sci 14: 1-5. |
| [38] | Liu F, Stützel H (2004) Biomass partitioning, specific leaf area, and water use efficiency of vegetable amaranth (Amaranthus spp.) in response to drought stress. Sci Hortic 102: 15-27. |
| [39] | Griñán I, Rodrgí uez P, Cruz ZN, et al. (2019) Leaf water relations in Diospyros kaki during a mild water deficit exposure. Agric Water Manage 217: 391-398. |
| [40] | Trabelsi L, Gargouri K, Ben Hassena A, et al. (2019) Impact of drought and salinity on olive water status and physiological performance in an arid climate. Agric Water Manage 213: 749-759. |
| [41] | Blanch M, Alvarez MD, Sanchez-Ballesta MT, et al. (2017) Water relations, short-chain oligosaccharides and rheological properties in lettuces subjected to limited water supply and low temperature stress. Sci Hortic 225: 726-735. |
| [42] | Ashraf M, Harris PJC (2013) Photosynthesis under stressful environments: An overview. Photosynthetica 51: 163-190. |
| [43] | Hernandez-Santana V, Fernández JE, Cuevas MV, et al. (2017) Photosynthetic limitations by water deficit: Effect on fruit and olive oil yield, leaf area and trunk diameter and its potential use to control vegetative growth of super-high density olive orchards. Agric Water Manage 184: 9-18. |
| [44] | Klamkowski K, Treder W (2008) Response to drought stress of three strawberry cultivars grown under greenhouse conditions. J Fruit Ornamental Plant Res 16: 179-188. |
| [45] | Liu B, Liang J, Tang G, et al. (2019) Drought stress effects on growth, water use efficiency, gas exchange and chlorophyll fluorescence of Juglans rootstocks. Sci Hortic 250: 230-235. |
| [46] | Srivastava LM (2002) CHAPTER 16-Abscisic Acid and Stress Tolerance in Plants. San Diego: Academic Press, 381-412. |
| [47] | Basu S, Ramegowda V, Kumar A, et al. (2016) Plant adaptation to drought stress. Research 5: 1554. |
| [48] |
Correia B, Pintó‐Marijuan M, Neves L, et al. (2014) Water stress and recovery in the performance of two Eucalyptus globulus clones: Physiological and biochemical profiles. Physiol Plant 150: 580-592. doi: 10.1111/ppl.12110
|
| [49] | Puangbut D, Jogloy S, Vorasoot N (2017) Association of photosynthetic traits with water use efficiency and SPAD chlorophyll meter reading of Jerusalem artichoke under drought conditions. Agric Water Manage 188: 29-35. |
| [50] |
Paknejad F, Nasri M, Moghadam HRT, et al. (2007) Effects of drought stress on chlorophyll fluorescence parameters, chlorophyll content and grain yield of wheat cultivars. J Biol Sci 7: 841-847. doi: 10.3923/jbs.2007.841.847
|
| [51] | Ju Y, Yue X, Zhao X, et al. (2018) Physiological, micro-morphological and metabolomic nalysis of grapevine (Vitis vinifera L.) leaf of plants under water stress. Plant Physiol Biochem 130: 501-510. |
| [52] | Naidoo G, Naidoo KK (2018) Drought stress effects on gas exchange and water relations of the invasive weed Chromolaena odorata. Flora 248: 1-9. |
| [53] |
Li Y, Li H, Li Y, et al. (2017) Improving water-use efficiency by decreasing stomatal conductance and transpiration rate to maintain higher ear photosynthetic rate in drought-resistant wheat. Crop J 5: 231-239. doi: 10.1016/j.cj.2017.01.001
|
| [54] | Zargar SM, Gupta N, Nazir M, et al. (2017) Impact of drought on photosynthesis: Molecular perspective. Plant Gene 11: 154-159. |
| [55] | Pessarakli M, Haghighi M, Sheibanirad A (2015) Plant responses under environmental stress conditions. Adv Plants Agric Res 2: 276-286. |
| [56] | Hussain M, Farooq S, Hasan W, et al. (2018) Drought stress in sunflower: Physiological effects and its management through breeding and agronomic alternatives. Agric Water Manage 201: 152-166. |
| [57] | Toscano S, Farieri E, Ferrante A, et al. (2016) Physiological and Biochemical Responsesin Two Ornamental Shrubs to Drought Stress. Front Plant Sci 7: 645. |
| [58] | Guo YY, Yu HY, Yang MM, et al. (2018) Effect of drought stress on lipid peroxidation, osmotic adjustment and antioxidant enzyme activity of leaves and roots of Lycium ruthenicum Murr. Seedling. Russ J Plant Physiol 65: 244-250. |
| [59] | Zhao XH, Yu HQ, Jing W, et al. (2016) Response of root morphology, physiology and endogenous hormones in maize (Zea mays L.) to potassium deficiency. J Integr Agric 15: 785-794. |
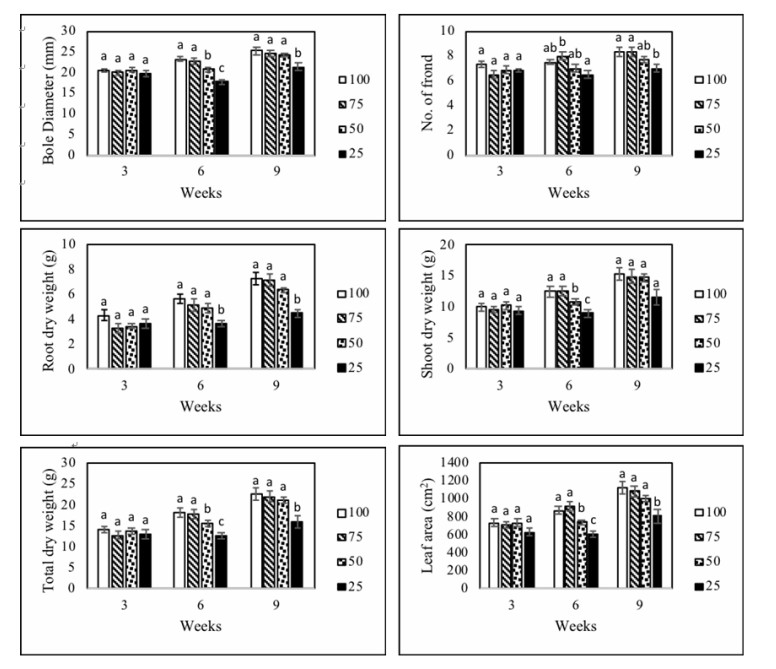
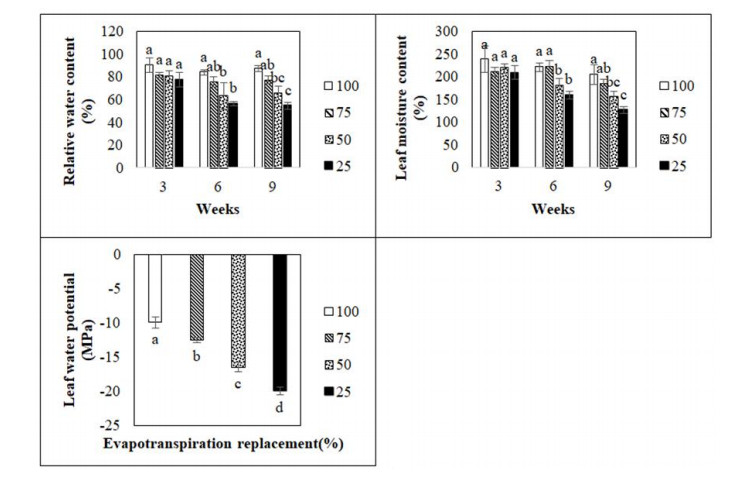
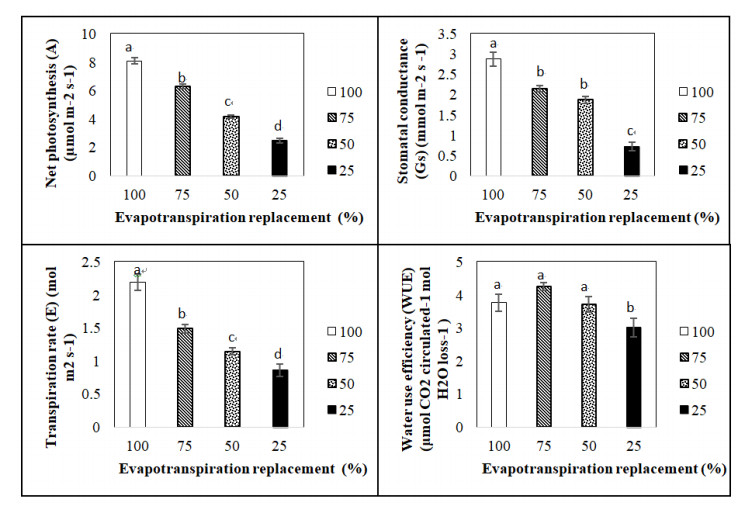
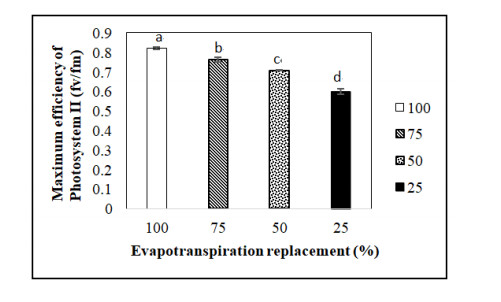
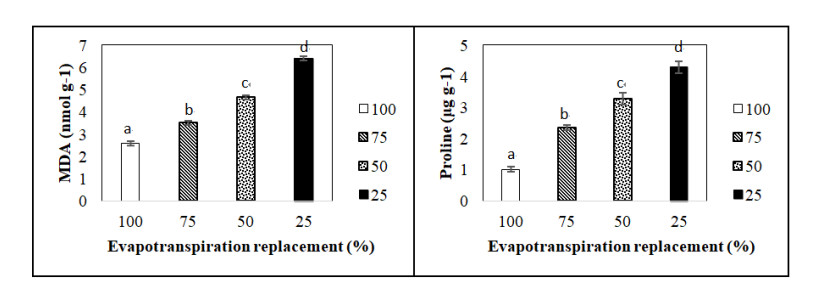
Figures(5) / Tables(1)

Tuan Syaripah Najihah, Mohd Hafiz Ibrahim, A'fifah Abd Razak, Rosimah Nulit, Puteri Edaroyati Megat Wahab. Effects of water stress on the growth, physiology and biochemical properties of oil palm seedlings[J]. AIMS Agriculture and Food, 2019, 4(4): 854-868. doi: 10.3934/agrfood.2019.4.854







 DownLoad:
DownLoad: