Citation: Md. Arman Arefin, Avijit Mallik. Numerical investigation of solar and wind energy assisted plugged in hybridized engine driven auto-rickshaw/three-wheeler[J]. AIMS Energy, 2018, 6(2): 358-375. doi: 10.3934/energy.2018.2.358

| [1] | Mehrdad E, Gao Y, Longo S, et al. (2018) Design and control methodology of series–parallel (torque and speed coupling) hybrid drivetrain, In: Mehrdad E, Gao Y, Longo S, et al. Modern electric, hybrid electric, and fuel cell vehicles, 10 Eds., CRC Press, 255–270. |
| [2] | Scott C, Robert MW, Russ C (2017) Summary report on the SAE 2016 range extenders for electric vehicles symposium. Available from: https://info.ornl.gov/sites/publications/files/Pub73742.pdf. |
| [3] |
Lane B, Brendan S, Samuelsen GS (2017) Plug-in fuel cell electric vehicles: A California case study. Int J Hydrogen Energ 42: 14294–14300. doi: 10.1016/j.ijhydene.2017.03.035

|
| [4] |
Ejaz S, Anwar K, Ashraf M, et al. (2009) Anti-angiogenic activities associated with exposure of environmental smoke solutions from 2-stroke auto-rickshaw. Environ Toxicol Phar 28: 42–51. doi: 10.1016/j.etap.2009.02.002
|
| [5] |
Mulhall P, Lukic SM, Wirasingha SG, et al. (2010) Solar-assisted electric auto rickshaw three-wheeler. IEEE T Veh Technol 59: 2298–2307. doi: 10.1109/TVT.2010.2045138
|
| [6] |
Kumarage, Amal S, Bandara M (2010) Analysis of the economic and social parameters of the Three-Wheeler Taxi service in Sri Lanka. Res Transp Econ 29: 395–400. doi: 10.1016/j.retrec.2010.07.050
|
| [7] | Chowdhury SJ, Rahman R, Azad A (2015) Power conversion for environment friendly electrically assisted rickshaw using photovoltaic technology in Bangladesh. Transp Electrification Conf Expo 2015: 1–6. |
| [8] |
Kelly JC, Macdonald JS, Keoleian GA (2012) Time-dependent plug-in hybrid electric vehicle charging based on national driving patterns and demographics. Appl Energ 94: 395–405. doi: 10.1016/j.apenergy.2012.02.001
|
| [9] | Mani A (2012) Sustainable urban transport in India: Role of the auto-rickshaw sector. Available from: http://www.indiaenvironmentportal.org.in/files/file/sustainable_urban_transport_india.pdf. |
| [10] | Siddique ARM, Sakib SN, Tahsien SM, et al. (2014) Performance evaluation of a microcontroller-based solar powered auto-rickshaw. Electr Eng Inf Commun Technol 2014: 1–6. |
| [11] | Shaha N, Uddin MB (2014) Hybrid energy assisted electric auto rickshaw three-wheeler. Electr Inform Commun Technol 7: 1–6. |
| [12] | Rashid MJA, Rahman A, Siddique ARM, et al. (2017) A novel approach of improving battery performance and longevity of the developed electrically assisted triwheeler vehicle by implementing torque sensor technology. Int J Veh Technol 2017: 1–16. |
| [13] |
Adak P, Sahu R, Elumalai SP (2016) Development of emission factors for motorcycles and shared auto-rickshaws using real-world driving cycle for a typical Indian city. Sci Total Environ 544: 299–308. doi: 10.1016/j.scitotenv.2015.11.099
|
| [14] | Arefin MA, Mallik A (2017) Solar assisted micro hybridized auto-rickshaw (engine driven). J Mech Eng Res Dev 40: 660–672. |
| [15] | Awal MR, Jusoh M, Sakib MN, et al. (2015) Design and implementation of vehicle mounted wind turbine. ARPN J Eng Appl Sci 10: 8699–8860. |
| [16] |
Chen Z, Xiong R, Wang K, et al. (2015) Optimal energy management strategy of a plug-in hybrid electric vehicle based on a particle swarm optimization algorithm. Energies 8: 3661–3678. doi: 10.3390/en8053661
|
| [17] | Serrao L, Onori S, Rizzoni G (2011) A comparative analysis of energy management strategies for hybrid electric vehicles. J Dyn Syst Meas Control 133: 1–9. |
| [18] |
Delgado J, Faria R, Moura P, et al. (2018) Impacts of plug-in electric vehicles in the portuguese electrical grid. Transp Res Part D 62: 372–385. doi: 10.1016/j.trd.2018.03.005
|
| [19] | Ferdous SM, Khaled WB, Ahmed B, et al. (2011) Electric vehicle with charging facility in motion using wind energy. World Renew Energ Congr-Sweden, 3629–3636 |
| [20] |
Fernandez-Gamiz U, Zulueta E, Boyano A, et al. (2017) Microtab design and implementation on a 5 MW wind turbine. Appl Sci 7: 536. doi: 10.3390/app7060536
|
| [21] | Masud MH, Naim-Ul-Hasan, Arefin AME, et al. (2017) Design modification of airfoil byintegrating sinusoidal leading edge and dimpled surface. AIP Conf Proc 1851: 191–209. |
| [22] |
Mulhall P, Lukic SM, Wirasingha SG, et al. (2010) Solar-assisted electric auto rickshaw three-wheeler. IEEE T Veh Technol 59: 2298–2307. doi: 10.1109/TVT.2010.2045138
|
| [23] |
Lukic SM, Mulhall P, Emadi A (2008) Energy autonomous solar/battery auto rickshaw. JAEV 6: 1135–1143. doi: 10.4130/jaev.6.1135
|
| [24] |
Fernandez-Gamiz U, Zulueta E, Boyano A, et al. (2017) Five megawatt wind turbine power output improvements by passive flow control devices. Energies 10: 742. doi: 10.3390/en10060742
|
| [25] | Joardder MUH (2013) Socio-economic & environmental impacts of battery driven auto rickshaw at Rajshahi city in Bangladesh. Int Conf Mech. |
| [26] | Furch J (2016) A model for predicting motor vehicle life cycle cost and its verification. Trans Famena 40: 15–26. |
| [27] |
Gransberg D (2010) Life cycle costing for engineers. Constr Manage Econ 28: 1113–1114. doi: 10.1080/01446193.2010.508500
|
| [28] | Mai ZQ, Xu TT, Sun L, et al. (2012) Cost estimation of product life cycle. Adv Mat Res 421: 582–585. |
| [29] |
Seo KK, Park JH, Jang DS, et al. (2002) Prediction of the life cycle cost using statistical and artificial neural network methods in conceptual product design. Int J Comput Integ Manuf 15: 541–554. doi: 10.1080/09511920210143417
|
| [30] |
Cai X, Tyagi S (2014) Development of a product life-cycle cost estimation model to support engineering decision-making in a multi-generational product development environment. J Cost Anal Parametrics 7: 219–235. doi: 10.1080/1941658X.2014.982403
|
| [31] |
Walker PD, Roser HM (2015) Energy consumption and cost analysis of hybrid electric powertrain configurations for two wheelers. Appl Energ 146: 279–287. doi: 10.1016/j.apenergy.2015.02.009
|
| [32] |
Requia WJ, Adams MD, Arain A, et al. (2017) Carbon dioxide emissions of plug-in hybrid electric vehicles: A life-cycle analysis in eight Canadian cities. Renew Sust Energ Rev 78: 1390–1396. doi: 10.1016/j.rser.2017.05.105
|
| [33] |
Rao AB, Rubin ES (2002) A technical, economic, and environmental assessment of amine-based CO2 capture technology for power plant greenhouse gas control. Environ Sci Technol 36: 4467–4475. doi: 10.1021/es0158861
|

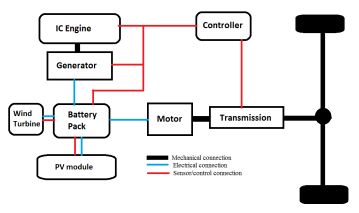
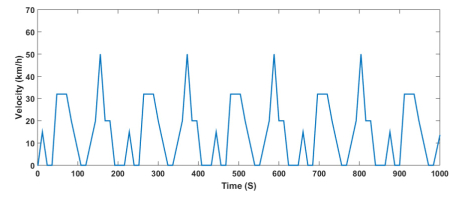
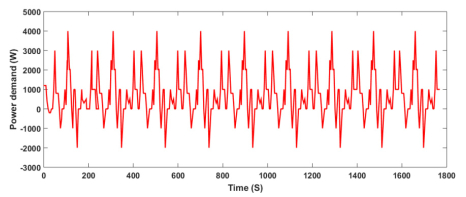
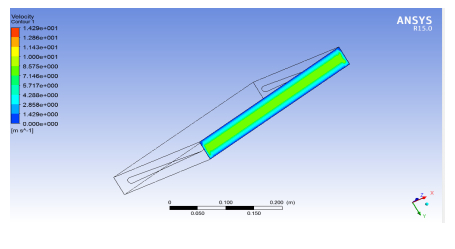
Figures(13) / Tables(9)

Md. Arman Arefin, Avijit Mallik. Numerical investigation of solar and wind energy assisted plugged in hybridized engine driven auto-rickshaw/three-wheeler[J]. AIMS Energy, 2018, 6(2): 358-375. doi: 10.3934/energy.2018.2.358







 DownLoad:
DownLoad: