Citation: Nick K. Olrichs, J. Bernd Helms. Novel insights into the function of the conserved domain of the CAP superfamily of proteins[J]. AIMS Biophysics, 2016, 3(2): 232-246. doi: 10.3934/biophy.2016.2.232
| [1] | Mathieu F. M. Cellier . Evolutionary analysis of Slc11 mechanism of proton-coupled metal-ion transmembrane import. AIMS Biophysics, 2016, 3(2): 286-318. doi: 10.3934/biophy.2016.2.286 |
| [2] | Eda Suku, Alejandro Giorgetti . Common evolutionary binding mode of rhodopsin-like GPCRs: Insights from structural bioinformatics. AIMS Biophysics, 2017, 4(4): 543-556. doi: 10.3934/biophy.2017.4.543 |
| [3] | Albert W. Pilkington IV, Justin Legleiter . Challenges in understanding the structure/activity relationship of Aβ oligomers. AIMS Biophysics, 2019, 6(1): 1-22. doi: 10.3934/biophy.2019.1.1 |
| [4] | Samantha B. Gacasan, Daniel L. Baker, Abby L. Parrill . G protein-coupled receptors: the evolution of structural insight. AIMS Biophysics, 2017, 4(3): 491-527. doi: 10.3934/biophy.2017.3.491 |
| [5] | Angel Rivera-Calzada, Andrés López-Perrote, Roberto Melero, Jasminka Boskovic, Hugo Muñoz-Hernández, Fabrizio Martino, Oscar Llorca . Structure and Assembly of the PI3K-like Protein Kinases (PIKKs) Revealed by Electron Microscopy. AIMS Biophysics, 2015, 2(2): 36-57. doi: 10.3934/biophy.2015.2.36 |
| [6] | Larisa I. Fedoreyeva, Boris F. Vanyushin, Ekaterina N. Baranova . Peptide AEDL alters chromatin conformation via histone binding. AIMS Biophysics, 2020, 7(1): 1-16. doi: 10.3934/biophy.2020001 |
| [7] | Mahmoud Abdallat, Abdallah Barjas Qaswal, Majed Eftaiha, Abdel Rahman Qamar, Qusai Alnajjar, Rawand Sallam, Lara Kollab, Mohammad Masa'deh, Anas Amayreh, Hiba Mihyar, Hesham Aboushakra, Bayan Alkelani, Rawan Owaimer, Mohannad Abd-Alhadi, Salwa Ireiqat, Fahed Turk, Ahmad Daoud, Bashar Darawsheh, Ahmad Hiasat, Majd Alhalaki, Shahem Abdallat, Salsabiela Bani Hamad, Rand Murshidi . A mathematical modeling of the mitochondrial proton leak via quantum tunneling. AIMS Biophysics, 2024, 11(2): 189-233. doi: 10.3934/biophy.2024012 |
| [8] | Judit Faus-Garriga, Isabel Novoa, Andrés Ozaita . mTOR signaling in proteostasis and its relevance to autism spectrum disorders. AIMS Biophysics, 2017, 4(1): 63-89. doi: 10.3934/biophy.2017.1.63 |
| [9] | Daniela Meleleo, Cesare Sblano . Influence of cholesterol on human calcitonin channel formation. Possible role of sterol as molecular chaperone. AIMS Biophysics, 2019, 6(1): 23-38. doi: 10.3934/biophy.2019.1.23 |
| [10] | Ateeq Al-Zahrani, Natasha Cant, Vassilis Kargas, Tracy Rimington, Luba Aleksandrov, John R. Riordan, Robert C. Ford . Structure of the cystic fibrosis transmembrane conductance regulator in the inward-facing conformation revealed by single particle electron microscopy. AIMS Biophysics, 2015, 2(2): 131-152. doi: 10.3934/biophy.2015.2.131 |
The CAP superfamily consists of proteins found in thousands of species across the entire biological kingdom [1]. In addition, CAP proteins are involved in a wide variety of functions, including fertilization, tumor suppression and immune modulation. Family members are characterized by the presence of a CAP domain, also referred to as the SCP (Sperm Coating Protein) domain, which encompasses a conserved tertiary structure: a unique α-β-α sandwich fold in which α-helices flank a central antiparallel β-sheet. Four highly conserved signature motifs have been recognized [1]. Outside these sequences there is little homology within the CAP domain between family members [1]. Most family members have an extension or an additional domain at the C-terminus of the CAP domain. Furthermore, all but one CAP protein family members possess a signal peptide for secretion to the extracellular space where they exert endo- or paracrine functions. Therefore, the large variety of additional domains is thought to be responsible for the remarkable evolutionary diversity in functions of CAP family members. For example, regulation of ion channels is a well-characterized function that has been attributed to the C-terminal domain present in the CRISP subfamily [2,3]. The role of the CAP domain and its conserved motifs has, however, largely remained elusive. Currently, some functions have been attributed to the CAP domain in certain species, such as protease activity in cone snail venom, sterol binding/transport in yeast, inhibition of integrin function, and scavenger of eicosanoids in saliva of blood-feeding insects [4,5,6,7]. However, none of these functions has been shown to be present in a wide range of superfamily members. The structurally highly conserved domain suggests a more common functionality throughout evolution.
Phylogenetic analysis of mammalian CAP proteins shows GAPR-1 to branch off early in evolution from the yeast PRY protein family and GAPR-1 might well be the first mammalian CAP protein in evolution [1]. GAPR-1 is the only mammalian CAP protein lacking a signal sequence and consists almost exclusively of a CAP domain without additional extensions [1,8,9]. Together, these features make GAPR-1 a suitable candidate to study the role of the conserved structural nature of the CAP domain.
GAPR-1 (also known as GLIPR-2 or C9orf19) acts as a negative regulator of autophagy [10], a cellular process involved in the degradation of cellular components via a lysosomal pathway. Autophagy plays an essential role in the immune system as it provides defense against infection, neurodegenerative disorders, cancer, and aging [11]. The protein exerts its negative regulatory function in autophagy by interacting with the essential autophagy-related protein Beclin 1 [10]. This results in the retention of Beclin 1 at the Golgi complex and inhibition of autophagy by preventing Beclin 1 to initiate autophagy. GAPR-1 is a small (17 kDa), highly positively charged (pI 9.4) peripheral membrane protein that contains an N-terminal myristoyl group [12], which localizes the protein to lipid-enriched microdomains at the cytosolic leaflet of the Golgi apparatus. GAPR-1 is highly expressed in immune-related tissues and cells, indicating that GAPR-1 plays a role in the mammalian innate immune system, analogous to PR-1 proteins in plants. Recently, GAPR-1 was shown to mediate interferon 1 signaling activation in response to Toll-like receptor 4 [13]. Additionally, GAPR-1 expression is vastly increased in renal proximal tubular cells during kidney fibrogenesis and stimulates epithelial to mesenchymal cell transition [14]. GAPR-1 has also been detected in extracellular vesicles secreted from prostate epithelial cells [15].
The molecular mechanisms of GAPR-1 have been extensively studied and provide interesting clues about how GAPR-1 exerts its biological function(s). GAPR-1 binds to liposomes containing negatively charged lipids [16]. It also interacts with Caveolin-1 on Golgi membranes [12] and together with N-terminal myristoylation [12] these signals are likely to be involved in the recruitment of GAPR-1 to raft-like lipid microdomains in the Golgi membrane. Another notable feature of GAPR-1 is that it has a strong tendency to form homodimers, both in vitro and in vivo (Figure 1) [8,9]. The protein also crystallizes as a dimer [9] and interaction with inositol-hexakisphosphate (IP6) induces dimerization with an alternative configuration in which one of the monomeric subunits of the crystallographic dimer is rotated by 28.5 degrees [17]. Membrane binding to liposomes also induces GAPR-1 homodimerization and as a result rapid tethering of the liposomes [17].
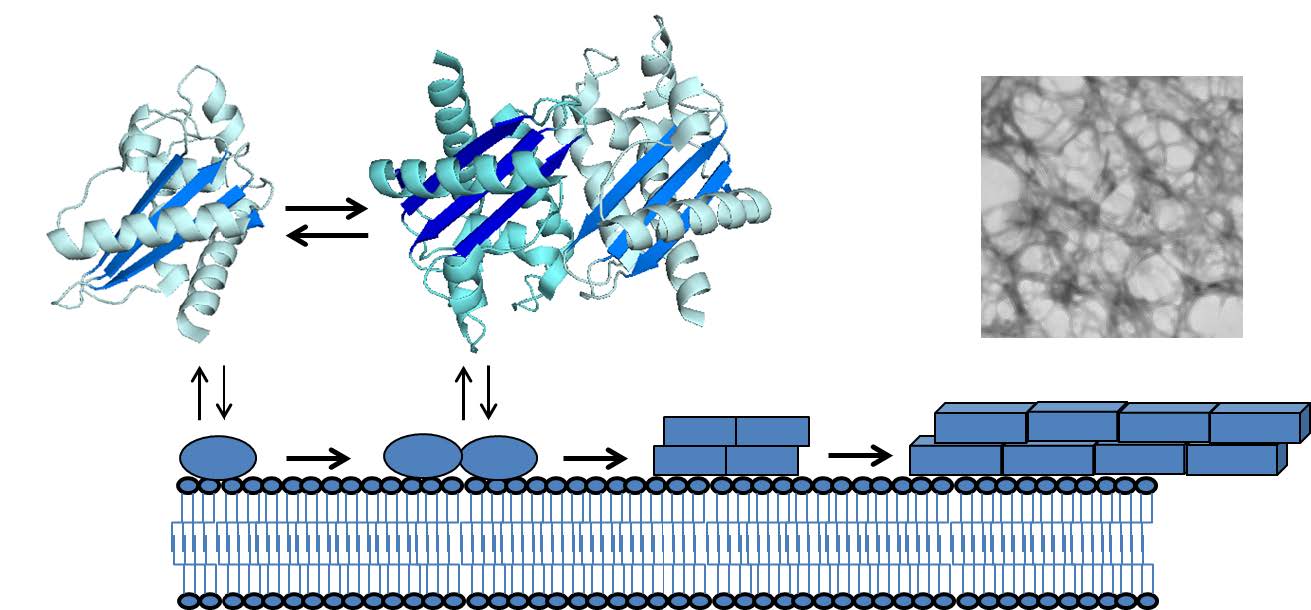 Figure 1. Schematic of GAPR-1 di-/oligomerization and membrane-induced amyloid formation. Monomer and dimer crystal structures (PDB: 1SMB)[9] and transmission EM image of GAPR-1 amyloid fibrils prepared as described in [18] are shown.
Figure 1. Schematic of GAPR-1 di-/oligomerization and membrane-induced amyloid formation. Monomer and dimer crystal structures (PDB: 1SMB)[9] and transmission EM image of GAPR-1 amyloid fibrils prepared as described in [18] are shown.
Prolonged incubation with negatively charged liposomes resulted in the formation of amyloid-like fibrils (Figure 1) [18]. The reasons for the propensity of GAPR-1 to form amyloid structures is not known, but it is intriguing to note that in the crystal structure of GAPR-1, the dimeric arrangement shows an almost continuous β-sheet extending beyond the monomeric units (Figure 1). The oligomerization process of GAPR-1 commences instantaneously upon membrane binding. The presence of cholesterol in the liposomes enhanced fibril formation, suggesting that localization of native GAPR-1 to lipid- and cholesterol-enriched microdomains of the Golgi complex favors oligomerization of GAPR-1 in the cell. Furthermore, natively folded GAPR-1 was shown to possess an intrinsic amyloid-related structure as it bound the amyloid oligomer-specific antibody A11 [18] and was shown to possess anti-β-amyloid aggregation activity [19]. GAPR-1 effectively inhibited Aβ fibril formation in vitro by binding to oligomeric Aβ structures [18]. The apparent paradoxical amyloid-forming and -inhibiting properties have been shown for several other proteins, including GroEL, small heat-shock proteins, α-synuclein and TTR [20,21,22]. GAPR-1 is also linked to amyloid-related diseases. In a study of necroptosis activation in multiple sclerosis (MS), GAPR-1 was found to be enriched in the insoluble proteome of MS patients [23]. GAPR-1 was also shown to be exclusively enriched in sites of neonatally induced neurodegeneration in rat hippocampus and its gene expression was proposed to regulate the development of diabetic neuropathy [24,25]. Whether these examples are related to the amyloid-related properties of GAPR-1, its role in autophagy regulation or both, remains to be investigated.
Self-association of proteins into dimers or oligomers can result in structural and functional benefits such as enhanced stability, control over active site accessibility and generation of new binding sites [26,27]. Oligomerization of proteins is known to be essential for a wide variety of processes, e.g. the regulation of its subcellular location, (de-)activation of enzymes and chaperones, regulation of gene expression and cell-cell adhesion [26,28]. Oligomerization can be mediated in several ways, including ligand binding, posttranslational modifications, disulfide bond formation between subunits and domain swapping. Oligomers serve diverse purposes in biological signaling to function as e.g. receptors or ligands, regulation of protein-protein interactions, or modulation of membrane curvature [26,29,30,31]. As both GAPR-1 and Beclin 1 form oligomeric structures [9,18,32,33], these properties may be intimately linked to regulate the interaction between these two proteins and/or regulate the activity of Beclin 1 in autophagy.
However, oligomerization can also lead to the formation of pathogenic protein structures. Proteins or peptides can undergo a specific self-assembly process into amyloid structures via a conserved pathway involving relatively short amino acid stretches that are normally hidden within the native structure of the protein, but upon exposure activate aggregation [34,35]. (Sub)cellular membranes and extracellular matrix components regularly play a critical role in initiating amyloid formation as they can serve as a structural template for self-assembly and often induce significant conformational changes in monomers or dimers leading to the exposure of amyloidogenic segments [36,37,38,39]. Subsequently, a conformational transition takes place from unstructured or native-like structures into oligomers rich in β-sheet structures. These oligomers act as seeds for adjacent monomers resulting in rapid elongation into mature amyloid fibrils, which are composed predominantly of highly ordered cross β-sheet structures [40]. Oligomerization of proteins and peptides into amyloid fibrils plays a central role in a wide variety of diseases, including Alzheimer’s, Parkinson’s and Huntington’s disease, prion disease and type II diabetes. Soluble amyloid oligomers are now recognized as the main toxic species [41]. However, the mechanisms of amyloid associated cytotoxicity are highly diverse and poorly understood [42]. One characteristic shared by most amyloid oligomers is their ability to disrupt membranes [43,44]. The membrane-perturbing property also renders amyloid peptides antimicrobial activity [45].
Intriguingly, there is a growing list of examples of amyloids serving a beneficial role for the organism. As short amyloid-forming peptides have been shown to possess catalytic activity in its fibrillar state, the amyloid fold has been hypothesized to be a common ancestral protein fold [46]. In contrast to disease-associated amyloids, the production of these so-called functional amyloids is tightly regulated without causing cytotoxicity. How this is achieved is not well understood. The regulation of protein abundance and fibrillation kinetics have been postulated as important factors in determining toxicity of amyloids [47]. Examples of functional amyloids include storage and release of hormones and toxins, serving as a template for pigment synthesis, and biofilm formation by bacteria and fungi [48,49]. A functional role for amyloids has also been implicated in maintenance of synaptic plasticity and memory consolidation [50,51], in cell adhesion [52], and as epigenetic elements of phenotypic inheritance [53]. Amyloid structures are also involved in signaling pathways. In programmed necrosis (necroptosis), the RIP1 and RIP3 proteins interact to form an amyloid core in the necrosome [54]. The functional amyloid NLRP3 inflammasome can be activated in response to amyloid fibrils from diverse sources, including Aβ, prions, serum amyloid A as well as curli from E. coli and S. Typhimurium [55]. Amyloid curli from these bacteria also activate Toll-like receptor 2 in intestinal epithelial cells, which leads to an enhanced barrier function and ameliorated inflammation [56]. Chronic activation of the immune system is now believed to be a major factor in neurodegenerative diseases and cardiac diseases [55,57].
The amylogenic properties of GAPR-1 and the emergence of functional amyloids early in evolution prompted us to consider the possibility that the CAP domain allows functional oligomeric and/or amyloidogenic regulation of the diverse CAP protein family members. Numerous CAP proteins have already been shown to exist and/or function as dimers or oligomers [9,58,59,60,61,62,63,64]. Furthermore, amyloid prediction analysis shows that CAP proteins in all taxa contain amyloidogenic segments within their CAP domain, with remarkably consistent motifs in the conserved CAP1 and CAP2 signature (Figure 2). Below we will discuss the indications for functional oligomeric and/or amylogenic interactions of CAP proteins in reproduction and immune regulation.
 Figure 2. Prediction of amyloidogenic regions in the CAP protein superfamily. CAP domain sequence alignment of CAP proteins from mammals, reptiles, insects, yeast and plants. The predicted amyloidogenic regions, obtained from amyloid prediction tool AmylPred2 [65] are highlighted in green. CAP signature motifs are boxed and indicated below the sequences. Modified from Olrichs et al. [18].
Figure 2. Prediction of amyloidogenic regions in the CAP protein superfamily. CAP domain sequence alignment of CAP proteins from mammals, reptiles, insects, yeast and plants. The predicted amyloidogenic regions, obtained from amyloid prediction tool AmylPred2 [65] are highlighted in green. CAP signature motifs are boxed and indicated below the sequences. Modified from Olrichs et al. [18].
Functional amyloids play a major role in fertilization. The zona pellucida (ZP), a glycoprotein extracellular matrix surrounding oocytes that is essential during fertilization, was recently discovered to possess all the classical characteristics of amyloid [66]. Functional amyloids are also present within the epididymal lumen and in the sperm acrosomal matrix, which interacts with ZP [67]. Also, mating of haploid yeast cells involves aggregation of cells promoted by amyloid-forming adhesion proteins [68,69]. Therefore, amyloidogenesis could be a conserved mechanism in reproduction throughout evolution.
A number of CAP proteins play pivotal roles in mammalian reproduction. These include mammalian sperm-binding proteins, CRISP1-4 and GLIPR1L1, which have been postulated to function in spermatogenesis, sperm capacitation and sperm-egg binding and fusion [70,71]. The CRISP subfamily is characterized by the presence of a cysteine-rich C-terminal extended domain, separated from the CAP domain by a hinge region [1]. The cysteine-rich C-terminal domain has ion channel regulatory activity, as was also determined for CRISP proteins derived from the venom of poisonous reptiles [2,3]. However, the role of the CAP domain remains unclear.
CRISP1 is essential for sperm maturation, capacitation and sperm-oocyte binding. It is expressed in the epididymis, secreted to the epididymal lumen and binds to spermatozoa in two populations: a loosely and tightly bound population. Zn2+binding was shown to induce formation of high molecular weight oligomeric CRISP1 complexes and facilitate its association to spermatozoa [58]. Following capacitation, the remaining tightly bound CRISP1 is involved in ZP and oocyte binding. For oocyte binding the CAP2 signature motif is essential. CRISP2 is a component of the sperm acrosome and following release during the acrosome reaction, it strongly binds to the sperm equatorial segment and is involved in sperm-egg fusion, similarly to and possibly in cooperation with CRISP1 [72]. CRISP3 and CRISP4 are also important for sperm-ZP binding. CRISP3 in equine seminal plasma was shown to suppress neutrophil-sperm binding, protecting sperm from elimination from the female reproductive tract [73]. GLIPR1L1 is localized at lipid rafts of spermatozoa and potentially also involved in ZP and oocyte binding [71].
CRISP1 is also present in the female genital tract. It is expressed by cumulus cells surrounding oocytes and capable of modulating sperm ion channels [74]. Allurin, a CRISP homolog from the female tract of Xenopus lacking the C-terminal CRISP domain was shown to function as a chemoattractant for sperm cells, also in mice [75,76]. Allurin forms SDS-insoluble oligomers typical of amyloid [62]. In relation to this, semen derived amyloid peptides were shown to enhance HIV infection by promoting virion fusion with host cells [77,78]. Due to the many commonalities between HIV infection and mammalian fertilization [79], in addition to the presence of amyloid proteins on the surface of spermatozoa and oocytes, amyloid fibrils are likely to play a role in sperm-egg binding and fusion. Recombinant human CRISP2 was already shown to have amyloid properties upon membrane binding in vitro [18]. Nevertheless, it remains to be established whether amyloidogenic and/or amyloid modulating properties are common to and important for the function of CAP proteins involved in fertilization.
Microbial pathogens of plants and mammals secrete an assortment of effector proteins that contribute to virulence by directly targeting host cells or tissues e.g. by suppressing defense responses. Evidence is mounting that amyloids play a role in evasion of immune response and host invasion [80]. Amyloid structures formed on the cell surfaces of bacteria and fungi not only function in microbial biofilm formation and cell adhesion, but were also shown to function as toxin storage and release sites and to inhibit neutrophil response by binding serum amyloid P [80,81,82]. Chronic infection with these pathogens has even been implicated as a causative factor in Alzheimer’s disease [83]. Harpin proteins secreted by various plant pathogens cause a hypersensitive response in the host plant (a process similar to apoptosis in mammalian cells), which was shown to be dependent on amyloid formation by the harpin [84].
In plants, the PR family of proteins have long been considered hallmarks of the hypersensitive response/defense pathways in plants. Among the 17 classified PR subfamilies [85], PR-1 is the only group which has not been attributed a biochemical function nor assigned any protein category with a recognized function. PR-1 proteins have been implicated in enhancing resistance against viral, fungal and oomycete infection [85,86,87]. There are various examples for PR-1 proteins from both host and pathogen where oligomerization seems to be crucial in the regulation of immunity, which will be discussed below. PR-1 proteins have been shown to accumulate at intercellular spaces between host and pathogen and to associate with fibrillary and electron-dense material [88,89,90]. PR-1 proteins were also found to localize in a clustered manner at the surface and in the cytoplasm of infection hyphae, inhibiting their differentiation [88,91]. The PR-1 protein homologue PR1-5 from hexaploid wheat was shown to physically interact with the Stagonospora nodorum toxin ToxA. This interaction potentially mediates toxin-induced necrosis in sensitive wheat. PR1-5 exists as a homodimer and mutation of the ToxA binding site did not affect dimerization. In a previous study the dimer conformation of PR1-5 was demonstrated to convey resistance to proteases [59]. PR-1 proteins were discovered that were fused via a transmembrane part to a kinase domain, reminiscent of receptor-like kinases [92]. These PR-1 receptor kinases were proposed to transduce defense signals through extracellular interaction with a PR-1 ligand. Ligand-induced oligomerization is a well-known mechanism for receptor kinase activation [93].
Recently, a wound induced peptide derived from the C-terminal part of PR-1, termed CAPE, was identified in tomato [94]. These peptides were shown to activate immune signals for anti-pathogen defense including its own precursor PR-1b. Homologous peptides in Arabidopsis were subsequently found to play an important role in the regulation of salt stress response [95]. Interestingly, the cleavage site of these peptides is before the proline residue within the CAP2 motif but after the predicted amyloidogenic part of the CAP2 motif, leaving the putative amyloidogenic properties intact (Figure 2).
Plant and mammalian pathogens also secrete PR-1-like proteins, which play an important role in pathogenicity. The fungus Fusarium oxysporum secretes a PR-1-like CAP protein, Fpr1, which was shown to be dispensable for virulence in plants but required for virulence in mammalian hosts [61]. Fpr1 forms dimers in solution and is proteolytically processed. As for PR-1-like proteins found in the saliva of blood-feeding insects, Fpr1 could be instrumental in evading the host immune system or to prevent blood clotting. Another possibility could be that fungi secrete PR-1-like proteins for antimicrobial actions against competing pathogens. This was proposed for a recently identified PR-1 protein with anti-bacterial activity in the digestive fluid of carnivorous plants [96].
PR-1-like proteins also have an important function in the infection process of parasitic nematodes through suppression and evasion of the host immune system. Upon host entry, they secrete a high amount of PR-1-like proteins, including a neutrophil inhibiting protein [97,98]. These ancylostoma secreted proteins (ASPs) are prime candidates for vaccine development. The human parasite Necator americanus produces two major ASPs that are thought to contribute to immune evasion and inhibition of platelet aggregation [63,99]. Intriguingly, Na-ASP-1 possesses two CAP domains that are linked via an extended loop [99]. Double domain ASPs are also capable of forming homodimers typical of the single-CAP domain ASPs [60,63]. Interestingly, the two-CAP domain protein was not effective in eliciting an immune response as opposed to single CAP ASPs, providing a clue as to how oligomerization of PR-1 proteins from other pathogens could play a part in immune evasion [99].
Other CAP protein family members have also been implicated in immune regulation. Hookworm platelet inhibitor (HPI) was discovered to inhibit platelet activation by blocking cell surface integrin receptors for fibrinogen and collagen [100]. HPI, like GAPR-1, crystallizes as a dimer although the orientation of the monomeric subunits is different as compared to dimeric GAPR-1 [64]. Remarkably, purified recombinant HPI was shown to be monomeric in solution, but unable to inhibit platelet adhesion to fibrinogen and collagen. Human neutrophil alpha defensins (HNPs) have been shown to bind the fibrinogen receptor and thereby induce formation of fibrinogen and thrombospondin-1 amyloid-like structures that activate platelets and bind microorganisms [101]. Amyloid peptides involved in Alzheimer’s disease are also intimately linked to platelet activation [102]. This could indicate that oligomeric HPI and/or amyloid-like fibrinogen are required for inhibition.
A similar example is natrin, a CRISP from snake venom, which functions as an inflammatory modulator by inducing expression of adhesion proteins in vascular endothelial cells. It was suggested that Zn2+ binding to the CAP domain in the presence of heparan sulfate enhances di-/oligomerization of natrin to activate this pathway [103]. Both zinc cations and heparan sulfate have been widely associated with amyloid formation [36,104,105].
Oligomerization and amyloid formation could be a more general mechanism in plant defense by other PR protein subfamilies as well, despite the fact that they do not contain the CAP domain. Prohevein is a wound induced antipathogenic protein from the rubber tree Hevea brasiliensis and the major allergen in latex. It is homologous to PR-4 proteins and has agglutination and chitinase properties. Its C-terminal domain, common to all PR-4 proteins, was shown to have amyloid-forming characteristics [106]. Other examples are found in PR-12 proteins (class II defensins), where dimer formation of tobacco NaD1 is critical for its antifungal activity [107]. Intriguingly, phosphatidylinositol binding of NaD1 induced its oligomerization and permeabilization of fungal and mammalian (tumor) cells. Nearly identical properties were revealed for tomato defensin TPP3, which formed amyloid-like fibrils upon specific binding to phosphatidylinositol (4, 5)-bisphosphate (PIP2) [108]. In radish seeds, a C-terminal peptide derived from antifungal defensins showed a high amyloid fibril-forming propensity [109]. The occurrence of haze during wine production has been attributed to the self-aggregation of PR proteins, such as thaumatin-like proteins (PR-5) and chitinases [110]. Besides defense against pathogens, PR proteins are also upregulated upon exposure to environmental stress. In winter rye, PR proteins from three different classes act synergistically as antifreeze proteins in oligomeric complexes [111].
CAP proteins are involved in a wide variety of biological functions. Yet, the function of the highly conserved CAP domain has not been elucidated. The structural properties and dynamics of the mammalian CAP protein GAPR-1 provide potential clues on this issue. We propose that oligomerization and amyloid formation/modulation is a common functionality of the CAP domain. There is evidence for a functional role of amyloid-forming proteins in cell-cell adhesion processes during fertilization, signaling pathways and immune regulation in host-pathogen interactions. Given the ubiquitous nature of amyloid folds in proteins throughout evolution, we suggest that the molecular mechanisms of GAPR-1 related to the structure of the CAP domain are characteristic for other members of the CAP superfamily of proteins as well.
This work was supported by National Institutes of Health grant U19AI109725 (J. B. H.).
All authors declare no conflicts of interest in this paper.
| [1] | Gibbs GM, Roelants K, O'Bryan MK (2008) The CAP superfamily: cysteine-rich secretory proteins, antigen 5, and pathogenesis-related 1 proteins--roles in reproduction, cancer, and immune defense. Endocr Rev 29: 865–897. |
| [2] | Gibbs GM, O'Bryan MK (2007) Cysteine rich secretory proteins in reproduction and venom. Soc Reprod Fertil Suppl 65: 261–267. |
| [3] | Yamazaki Y, Morita T (2004) Structure and function of snake venom cysteine-rich secretory proteins. Toxicon 44: 227–231. |
| [4] | Choudhary V, Schneiter R (2012) Pathogen-Related Yeast (PRY) proteins and members of the CAP superfamily are secreted sterol-binding proteins. Proc Natl Acad Sci USA 109: 16882–16887. |
| [5] | Xu X, Francischetti IM, Lai R, et al. (2012) Structure of protein having inhibitory disintegrin and leukotriene scavenging functions contained in single domain. J Biol Chem 287: 10967–10976. |
| [6] | Kelleher A, Darwiche R, Rezende WC, et al. (2014) Schistosoma mansoni venom allergen-like protein 4 (SmVAL4) is a novel lipid-binding SCP/TAPS protein that lacks the prototypical CAP motifs. Acta Crystallogr D Biol Crystallogr 70: 2186–2196. |
| [7] | Milne TJ, Abbenante G, Tyndall JD, et al. (2003) Isolation and characterization of a cone snail protease with homology to CRISP proteins of the pathogenesis-related protein superfamily. J Biol Chem 278: 31105–31110. |
| [8] | Eberle HB, Serrano RL, Fullekrug J, et al. (2002) Identification and characterization of a novel human plant pathogenesis-related protein that localizes to lipid-enriched microdomains in the Golgi complex. J Cell Sci 115: 827–838. |
| [9] | Serrano RL, Kuhn A, Hendricks A, et al. (2004) Structural analysis of the human Golgi-associated plant pathogenesis related protein GAPR-1 implicates dimerization as a regulatory mechanism. J Mol Biol 339: 173–183. |
| [10] | Shoji-Kawata S, Sumpter R, Leveno M, et al. (2013) Identification of a candidate therapeutic autophagy-inducing peptide. Nature 494: 201–206. |
| [11] | Kuballa P, Nolte WM, Castoreno AB, et al. (2012) Autophagy and the immune system. Annu Rev Immunol 30: 611–646. |
| [12] | Eberle HB, Serrano RL, Fullekrug J, et al. (2002) Identification and characterization of a novel human plant pathogenesis-related protein that localizes to lipid-enriched microdomains in the Golgi complex. J Cell Sci 115: 827–838. |
| [13] | Zhou Q, Hao L, Huang W, et al. (2016) The Golgi-Associated Plant Pathogenesis-Related Protein GAPR-1 Enhances Type I Interferon Signaling Pathway in Response to Toll-Like Receptor 4. Inflammation 39: 706–717. |
| [14] | Baxter RM, Crowell TP, George JA, et al. (2007) The plant pathogenesis related protein GLIPR-2 is highly expressed in fibrotic kidney and promotes epithelial to mesenchymal transition in vitro. Matrix Biol 26: 20–29. |
| [15] | Aalberts M, van Dissel-Emiliani FM, van Adrichem NP, et al. (2012) Identification of distinct populations of prostasomes that differentially express prostate stem cell antigen, annexin A1, and GLIPR2 in humans. Biol Reprod 86: 82:1–8. |
| [16] | Van Galen J, Van Balkom BW, Serrano RL, et al. (2010) Binding of GAPR-1 to negatively charged phospholipid membranes: unusual binding characteristics to phosphatidylinositol. Mol Membr Biol 27: 81–91. |
| [17] | van Galen J, Olrichs NK, Schouten A, et al. (2012) Interaction of GAPR-1 with lipid bilayers is regulated by alternative homodimerization. Biochim Biophys Acta 1818: 2175–2183. |
| [18] | Olrichs NK, Mahalka AK, Kaloyanova D, et al. (2014) Golgi-Associated plant Pathogenesis Related protein 1 (GAPR-1) forms amyloid-like fibrils by interaction with acidic phospholipids and inhibits Abeta aggregation. Amyloid 21: 88–96. |
| [19] | Yoshiike Y, Minai R, Matsuo Y, et al. (2008) Amyloid oligomer conformation in a group of natively folded proteins. PLoS One 3: e3235. |
| [20] | Schwarzman AL, Gregori L, Vitek MP, et al. (1994) Transthyretin sequesters amyloid beta protein and prevents amyloid formation. Proc Natl Acad Sci USA 91: 8368–8372. |
| [21] | Chen J, Yagi H, Sormanni P, et al. (2012) Fibrillogenic propensity of the GroEL apical domain: a Janus-faced minichaperone. FEBS Lett 586: 1120–1127. |
| [22] | Rekas A, Ahn KJ, Kim J, et al. (2012) The chaperone activity of alpha-synuclein: Utilizing deletion mutants to map its interaction with target proteins. Proteins 80: 1316–1325. |
| [23] | Ofengeim D, Ito Y, Najafov A, et al. (2015) Activation of necroptosis in multiple sclerosis. Cell Rep 10: 1836–1849. |
| [24] | Zhang L, Qu S, Liang A, et al. (2015) Gene expression microarray analysis of the sciatic nerve of mice with diabetic neuropathy. Int J Mol Med 35: 333–339. |
| [25] | Karlsson O, Berg AL, Hanrieder J, et al. (2015) Intracellular fibril formation, calcification, and enrichment of chaperones, cytoskeletal, and intermediate filament proteins in the adult hippocampus CA1 following neonatal exposure to the nonprotein amino acid BMAA. Arch Toxicol 89: 423–436. |
| [26] | Nishi H, Hashimoto K, Madej T, et al. (2013) Evolutionary, physicochemical, and functional mechanisms of protein homooligomerization. Prog Mol Biol Transl Sci 117: 3–24. |
| [27] | Marianayagam NJ, Sunde M, Matthews JM (2004) The power of two: protein dimerization in biology. Trends Biochem Sci 29: 618–625. |
| [28] | Ali MH, Imperiali B (2005) Protein oligomerization: how and why. Bioorg Med Chem 13: 5013–5020. |
| [29] | Wang X, Sharp JS, Handel TM, et al. (2013) Chemokine oligomerization in cell signaling and migration. Prog Mol Biol Transl Sci 117: 531–578. |
| [30] | Zimmerberg J, Kozlov MM (2006) How proteins produce cellular membrane curvature. Nat Rev Mol Cell Biol 7: 9–19. |
| [31] | Park PS, Filipek S, Wells JW, et al. (2004) Oligomerization of G protein-coupled receptors: past, present, and future. Biochemistry 43: 15643–15656. |
| [32] | Adi-Harel S, Erlich S, Schmukler E, et al. (2010) Beclin 1 self-association is independent of autophagy induction by amino acid deprivation and rapamycin treatment. J Cell Biochem 110: 1262–1271. |
| [33] | Ku B, Woo JS, Liang C, et al. (2008) An insight into the mechanistic role of Beclin 1 and its inhibition by prosurvival Bcl-2 family proteins. Autophagy 4: 519–520. |
| [34] | Chiti F, Dobson CM (2006) Protein misfolding, functional amyloid, and human disease. Annu Rev Biochem 75: 333–366. |
| [35] | Dobson CM (2003) Protein folding and misfolding. Nature 426: 884–890. |
| [36] | Iannuzzi C, Irace G, Sirangelo I (2015) The effect of glycosaminoglycans (GAGs) on amyloid aggregation and toxicity. Molecules 20: 2510–2528. |
| [37] | Oskarsson ME, Singh K, Wang J, et al. (2015) Heparan Sulfate Proteoglycans Are Important for Islet Amyloid Formation and Islet Amyloid Polypeptide-induced Apoptosis. J Biol Chem 290: 15121–15132. |
| [38] | Kinnunen PK (2009) Amyloid Formation on Lipid Membrane Surfaces. The Open Biology Journal 2: 163–175. |
| [39] | Burke KA, Yates EA, Legleiter J (2013) Biophysical insights into how surfaces, including lipid membranes, modulate protein aggregation related to neurodegeneration. Front Neurol 4: 17. |
| [40] | Sunde M, Serpell LC, Bartlam M, et al. (1997) Common core structure of amyloid fibrils by synchrotron X-ray diffraction. J Mol Biol 273: 729–739. |
| [41] | Glabe CG (2008) Structural classification of toxic amyloid oligomers. J Biol Chem 283: 29639–29643. |
| [42] | Kayed R, Lasagna-Reeves CA (2013) Molecular mechanisms of amyloid oligomers toxicity. J Alzheimers Dis 33 Suppl 1: S67–78. |
| [43] | Breydo L, Uversky VN (2015) Structural, morphological, and functional diversity of amyloid oligomers. FEBS Lett 589: 2640–2648. |
| [44] | Haass C, Selkoe DJ (2007) Soluble protein oligomers in neurodegeneration: lessons from the Alzheimer's amyloid beta-peptide. Nat Rev Mol Cell Biol 8: 101–112. |
| [45] | Kagan BL, Jang H, Capone R, et al. (2012) Antimicrobial properties of amyloid peptides. Mol Pharm 9: 708–717. |
| [46] | Greenwald J, Riek R (2012) On the possible amyloid origin of protein folds. J Mol Biol 421: 417–426. |
| [47] | Gsponer J, Babu MM (2012) Cellular strategies for regulating functional and nonfunctional protein aggregation. Cell Rep 2: 1425–1437. |
| [48] | Fowler DM, Koulov AV, Balch WE, et al. (2007) Functional amyloid--from bacteria to humans. Trends Biochem Sci 32: 217–224. |
| [49] | Maji SK, Perrin MH, Sawaya MR, et al. (2009) Functional amyloids as natural storage of peptide hormones in pituitary secretory granules. Science 325: 328–332. |
| [50] | Puzzo D, Arancio O (2013) Amyloid-beta peptide: Dr. Jekyll or Mr. Hyde? J Alzheimers Dis 33 Suppl 1: S111–20. |
| [51] | Hervas R, Li L, Majumdar A, et al. (2016) Molecular Basis of Orb2 Amyloidogenesis and Blockade of Memory Consolidation. PLoS Biol 14: e1002361. |
| [52] | Jacob RS, George E, Singh PK, et al. (2016) Cell Adhesion on Amyloid Fibrils Lacking Integrin Recognition Motif. J Biol Chem . |
| [53] | Halfmann R, Jarosz DF, Jones SK, et al. (2012) Prions are a common mechanism for phenotypic inheritance in wild yeasts. Nature 482: 363–368. |
| [54] | Li J, McQuade T, Siemer AB, et al. (2012) The RIP1/RIP3 necrosome forms a functional amyloid signaling complex required for programmed necrosis. Cell 150: 339–350. |
| [55] | Parry TL, Melehani JH, Ranek MJ, et al. (2015) Functional Amyloid Signaling via the Inflammasome, Necrosome, and Signalosome: New Therapeutic Targets in Heart Failure. Front Cardiovasc Med 2: 25. |
| [56] | Rapsinski GJ, Wynosky-Dolfi MA, Oppong GO, et al. (2015) Toll-like receptor 2 and NLRP3 cooperate to recognize a functional bacterial amyloid, curli. Infect Immun 83: 693–701. |
| [57] | Heneka MT, Kummer MP, Latz E (2014) Innate immune activation in neurodegenerative disease. Nat Rev Immunol 14: 463–477. |
| [58] | Maldera JA, Vasen G, Ernesto JI, et al. (2011) Evidence for the involvement of zinc in the association of CRISP1 with rat sperm during epididymal maturation. Biol Reprod 85: 503–510. |
| [59] | Lu S, Faris JD, Sherwood R, et al. (2014) A dimeric PR-1-type pathogenesis-related protein interacts with ToxA and potentially mediates ToxA-induced necrosis in sensitive wheat. Mol Plant Pathol 15: 650–663. |
| [60] | Borloo J, Geldhof P, Peelaers I, et al. (2013) Structure of Ostertagia ostertagi ASP-1: insights into disulfide-mediated cyclization and dimerization. Acta Crystallogr D Biol Crystallogr 69: 493–503. |
| [61] | Prados-Rosales RC, Roldan-Rodriguez R, Serena C, et al. (2012) A PR-1-like protein of Fusarium oxysporum functions in virulence on mammalian hosts. J Biol Chem 287: 21970–21979. |
| [62] | Sugiyama H, Burnett L, Xiang X, et al. (2009) Purification and multimer formation of allurin, a sperm chemoattractant from Xenopus laevis egg jelly. Mol Reprod Dev 76: 527–536. |
| [63] | Asojo OA, Goud G, Dhar K, et al. (2005) X-ray structure of Na-ASP-2, a pathogenesis-related-1 protein from the nematode parasite, Necator americanus, and a vaccine antigen for human hookworm infection. J Mol Biol 346: 801–814. |
| [64] | Ma D, Francischetti IM, Ribeiro JM, et al. (2015) The structure of hookworm platelet inhibitor (HPI), a CAP superfamily member from Ancylostoma caninum. Acta Crystallogr F Struct Biol Commun 71: 643–649. |
| [65] | Tsolis AC, Papandreou NC, Iconomidou VA, et al. (2013) A consensus method for the prediction of 'aggregation-prone' peptides in globular proteins. PLoS One 8: e54175. |
| [66] | Egge N, Muthusubramanian A, Cornwall GA (2015) Amyloid properties of the mouse egg zona pellucida. PLoS One 10: e0129907. |
| [67] | Guyonnet B, Egge N, Cornwall GA (2014) Functional amyloids in the mouse sperm acrosome. Mol Cell Biol 34: 2624–2634. |
| [68] | Swanson WJ, Aagaard JE, Vacquier VD, et al. (2011) The molecular basis of sex: linking yeast to human. Mol Biol Evol 28: 1963–1966. |
| [69] | Garcia MC, Lee JT, Ramsook CB, et al. (2011) A role for amyloid in cell aggregation and biofilm formation. PLoS One 6: e17632. |
| [70] | Da Ros VG, Munoz MW, Battistone MA, et al. (2015) From the epididymis to the egg: participation of CRISP proteins in mammalian fertilization. Asian J Androl 17: 711–715. |
| [71] | Caballero J, Frenette G, D'Amours O, et al. (2012) Bovine sperm raft membrane associated Glioma Pathogenesis-Related 1-like protein 1 (GliPr1L1) is modified during the epididymal transit and is potentially involved in sperm binding to the zona pellucida. J Cell Physiol 227: 3876–3886. |
| [72] | Busso D, Goldweic NM, Hayashi M, et al. (2007) Evidence for the involvement of testicular protein CRISP2 in mouse sperm-egg fusion. Biol Reprod 76: 701–708. |
| [73] | Doty A, Buhi WC, Benson S, et al. (2011) Equine CRISP3 modulates interaction between spermatozoa and polymorphonuclear neutrophils. Biol Reprod 85: 157–164. |
| [74] | Ernesto JI, Weigel Munoz M, Battistone MA, et al. (2015) CRISP1 as a novel CatSper regulator that modulates sperm motility and orientation during fertilization. J Cell Biol 210: 1213–1224. |
| [75] | Burnett LA, Xiang X, Bieber AL, et al. (2008) Crisp proteins and sperm chemotaxis: discovery in amphibians and explorations in mammals. Int J Dev Biol 52: 489–501. |
| [76] | Burnett LA, Anderson DM, Rawls A, et al. (2011) Mouse sperm exhibit chemotaxis to allurin, a truncated member of the cysteine-rich secretory protein family. Dev Biol 360: 318–328. |
| [77] | Usmani SM, Zirafi O, Muller JA, et al. (2014) Direct visualization of HIV-enhancing endogenous amyloid fibrils in human semen. Nat Commun 5: 3508. |
| [78] | Castellano LM, Shorter J (2012) The Surprising Role of Amyloid Fibrils in HIV Infection. Biology (Basel) 1: 58–80. |
| [79] | Doncel GF (2006) Exploiting common targets in human fertilization and HIV infection: development of novel contraceptive microbicides. Hum Reprod Update 12: 103–117. |
| [80] | Gebbink MF, Claessen D, Bouma B, et al. (2005) Amyloids--a functional coat for microorganisms. Nat Rev Microbiol 3: 333–341. |
| [81] | Garcia-Sherman MC, Lundberg T, Sobonya RE, et al. (2015) A unique biofilm in human deep mycoses: fungal amyloid is bound by host serum amyloid P component. NPJ Biofilms Microbiomes 1: 15009. |
| [82] | Syed AK, Boles BR (2014) Fold modulating function: bacterial toxins to functional amyloids. Front Microbiol 5: 401. |
| [83] | Pisa D, Alonso R, Rabano A, et al. (2015) Different Brain Regions are Infected with Fungi in Alzheimer's Disease. Sci Rep 5: 15015. |
| [84] | Oh J, Kim JG, Jeon E, et al. (2007) Amyloidogenesis of type III-dependent harpins from plant pathogenic bacteria. J Biol Chem 282: 13601–13609. |
| [85] | van Loon LC, Rep M, Pieterse CM (2006) Significance of inducible defense-related proteins in infected plants. Annu Rev Phytopathol 44: 135–162. |
| [86] | Van Loon LC, Van Strien EA (1999) The families of pathogenesis-related proteins, their activities, and comparative analysis of PR-1 type proteins. Physiol Mol Plant Pathol 55: 85–97. |
| [87] | Niderman T, Genetet I, Bruyere T, et al. (1995) Pathogenesis-related PR-1 proteins are antifungal. Isolation and characterization of three 14-kilodalton proteins of tomato and of a basic PR-1 of tobacco with inhibitory activity against Phytophthora infestans. Plant Physiol 108: 17–27. |
| [88] | Rauscher M, Adam AL, Wirtz S, et al. (1999) PR-1 protein inhibits the differentiation of rust infection hyphae in leaves of acquired resistant broad bean. Plant J 19: 625–633. |
| [89] | Benhamou N (1995) Immunocytochemistry of plant defense mechanisms induced upon microbial attack. Microsc Res Tech 31: 63–78. |
| [90] | Lee YK, Hippe-Sanwald S, Lee SC, et al. (2000) In situ localization of PR-1 mRNA and PR-1 protein in compatible and incompatible interactions of pepper stems with Phytophthora capsici. Protoplasma 211: 64–75. |
| [91] | Santén K, Marttila S, Liljeroth E, et al. (2005) Immunocytochemical localization of the pathogenesis-related PR-1 protein in barley leaves after infection by Bipolaris sorokiniana. Physiol Mol Plant Pathol 66: 45–54. |
| [92] | Teixeira PJ, Costa GG, Fiorin GL, et al. (2013) Novel receptor-like kinases in cacao contain PR-1 extracellular domains. Mol Plant Pathol 14: 602–609. |
| [93] | Jiang G, Hunter T (1999) Receptor signaling: when dimerization is not enough. Curr Biol 9: R568–71. |
| [94] | Chen YL, Lee CY, Cheng KT, et al. (2014) Quantitative peptidomics study reveals that a wound-induced peptide from PR-1 regulates immune signaling in tomato. Plant Cell 26: 4135–4148. |
| [95] | Chien PS, Nam HG, Chen YR (2015) A salt-regulated peptide derived from the CAP superfamily protein negatively regulates salt-stress tolerance in Arabidopsis. J Exp Bot 66: 5301–5313. |
| [96] | Buch F, Pauchet Y, Rott M, et al. (2014) Characterization and heterologous expression of a PR-1 protein from traps of the carnivorous plant Nepenthes mirabilis. Phytochemistry 100: 43–50. |
| [97] | Osman A, Wang CK, Winter A, et al. (2012) Hookworm SCP/TAPS protein structure--A key to understanding host-parasite interactions and developing new interventions. Biotechnol Adv 30: 652–657. |
| [98] | Moyle M, Foster DL, McGrath DE, et al. (1994) A hookworm glycoprotein that inhibits neutrophil function is a ligand of the integrin CD11b/CD18. J Biol Chem 269: 10008–10015. |
| [99] | Asojo OA (2011) Structure of a two-CAP-domain protein from the human hookworm parasite Necator americanus. Acta Crystallogr D Biol Crystallogr 67: 455–462. |
| [100] | Del Valle A, Jones BF, Harrison LM, et al. (2003) Isolation and molecular cloning of a secreted hookworm platelet inhibitor from adult Ancylostoma caninum. Mol Biochem Parasitol 129: 167–177. |
| [101] | Horn M, Bertling A, Brodde MF, et al. (2012) Human neutrophil alpha-defensins induce formation of fibrinogen and thrombospondin-1 amyloid-like structures and activate platelets via glycoprotein IIb/IIIa. J Thromb Haemost 10: 647–661. |
| [102] | Canobbio I, Abubaker AA, Visconte C, et al. (2015) Role of amyloid peptides in vascular dysfunction and platelet dysregulation in Alzheimer's disease. Front Cell Neurosci 9: 65. |
| [103] | Wang YL, Kuo JH, Lee SC, et al. (2010) Cobra CRISP functions as an inflammatory modulator via a novel Zn2+- and heparan sulfate-dependent transcriptional regulation of endothelial cell adhesion molecules. J Biol Chem 285: 37872–37883. |
| [104] | Alexandrescu AT (2005) Amyloid accomplices and enforcers. Protein Sci 14: 1–12. |
| [105] | Hughes AJ, Hussain R, Cosentino C, et al. (2012) A zinc complex of heparan sulfate destabilises lysozyme and alters its conformation. Biochem Biophys Res Commun 425: 794–799. |
| [106] | Berthelot K, Lecomte S, Coulary-Salin B, et al. (2016) Hevea brasiliensis prohevein possesses a conserved C-terminal domain with amyloid-like properties in vitro. Biochim Biophys Acta 1864: 388–399. |
| [107] | Lay FT, Mills GD, Poon IK, et al. (2012) Dimerization of plant defensin NaD1 enhances its antifungal activity. J Biol Chem 287: 19961–19972. |
| [108] | Poon IK, Baxter AA, Lay FT, et al. (2014) Phosphoinositide-mediated oligomerization of a defensin induces cell lysis. Elife 3: e01808. |
| [109] | Garvey M, Meehan S, Gras SL, et al. (2013) A radish seed antifungal peptide with a high amyloid fibril-forming propensity. Biochim Biophys Acta 1834: 1615–1623. |
| [110] | Marangon M, Van Sluyter SC, Waters EJ, et al. (2014) Structure of haze forming proteins in white wines: Vitis vinifera thaumatin-like proteins. PLoS One 9: e113757. |
| [111] | Yu XM, Griffith M (1999) Antifreeze proteins in winter rye leaves form oligomeric complexes. Plant Physiol 119: 1361–1370. |
| 1. | Yue Li, Yuting Zhao, Minfei Su, Karen Glover, Srinivas Chakravarthy, Christopher L. Colbert, Beth Levine, Sangita C. Sinha, Structural insights into the interaction of the conserved mammalian proteins GAPR-1 and Beclin 1, a key autophagy protein, 2017, 73, 2059-7983, 775, 10.1107/S2059798317011822 | |
| 2. | Avinash S Gaikwad, Jinghua Hu, David G Chapple, Moira K O’Bryan, The functions of CAP superfamily proteins in mammalian fertility and disease, 2020, 26, 1355-4786, 689, 10.1093/humupd/dmaa016 | |
| 3. | Marco Gerdol, Manuela Cervelli, Paolo Mariottini, Marco Oliverio, Sébastien Dutertre, Maria Modica, A Recurrent Motif: Diversity and Evolution of ShKT Domain Containing Proteins in the Vampire Snail Cumia reticulata, 2019, 11, 2072-6651, 106, 10.3390/toxins11020106 | |
| 4. | Jie Sheng, Nick K. Olrichs, Willie J. Geerts, Dora V. Kaloyanova, J. Bernd Helms, Metal ions and redox balance regulate distinct amyloid-like aggregation pathways of GAPR-1, 2019, 9, 2045-2322, 10.1038/s41598-019-51232-7 | |
| 5. | Nikolett Lupsa, Barbara Érsek, Andor Horváth, András Bencsik, Eszter Lajkó, Pálma Silló, Ádám Oszvald, Zoltán Wiener, Péter Reményi, Gábor Mikala, Tamás Masszi, Edit I Buzás, Zoltán Pós, Skin‐homing CD8 + T cells preferentially express GPI‐anchored peptidase inhibitor 16, an inhibitor of cathepsin K , 2018, 48, 0014-2980, 1944, 10.1002/eji.201847552 | |
| 6. | Jie Sheng, Nick K. Olrichs, Bart M. Gadella, Dora V. Kaloyanova, J. Bernd Helms, Regulation of Functional Protein Aggregation by Multiple Factors: Implications for the Amyloidogenic Behavior of the CAP Superfamily Proteins, 2020, 21, 1422-0067, 6530, 10.3390/ijms21186530 | |
| 7. | Jie Sheng, Bart M. Gadella, Nick K. Olrichs, Dora V. Kaloyanova, J. Bernd Helms, The less conserved metal-binding site in human CRISP1 remains sensitive to zinc ions to permit protein oligomerization, 2021, 11, 2045-2322, 10.1038/s41598-021-84926-y | |
| 8. | Jie Sheng, Nick K. Olrichs, Willie J. Geerts, Xueyi Li, Ashfaq Ur Rehman, Barend M. Gadella, Dora V. Kaloyanova, J. Bernd Helms, Zinc binding regulates amyloid-like aggregation of GAPR-1, 2019, 39, 0144-8463, 10.1042/BSR20182345 | |
| 9. | Yan Sun, Minxiao Wang, Hao Chen, Hao Wang, Zhaoshan Zhong, Li Zhou, Lulu Fu, Chaolun Li, Song Sun, Insights into symbiotic interactions from metatranscriptome analysis of deep‐sea mussel Gigantidas platifrons under long‐term laboratory maintenance , 2023, 32, 0962-1083, 444, 10.1111/mec.16765 | |
| 10. | Nafiseh Sirati, Blagovesta Popova, Martijn R. Molenaar, Iris C. Verhoek, Gerhard H. Braus, Dora V. Kaloyanova, J. Bernd Helms, Dynamic and Reversible Aggregation of the Human CAP Superfamily Member GAPR-1 in Protein Inclusions in Saccharomyces cerevisiae, 2021, 433, 00222836, 167162, 10.1016/j.jmb.2021.167162 | |
| 11. | Nafiseh Sirati, Ziying Shen, Nick K. Olrichs, Blagovesta Popova, Iris C. Verhoek, Ilse M. Lagerwaard, Gerhard H. Braus, Dora V. Kaloyanova, J.Bernd Helms, GAPR-1 Interferes with Condensate Formation of Beclin 1 in Saccharomyces cerevisiae, 2023, 435, 00222836, 167935, 10.1016/j.jmb.2022.167935 | |
| 12. | Qianqian Zhang, Jiawei Xu, Xi Zhou, Zhonghua Liu, CAP superfamily proteins from venomous animals: Who we are and what to do?, 2022, 221, 01418130, 691, 10.1016/j.ijbiomac.2022.09.079 | |
| 13. | Pengliang Han, Chengli Wang, Fudong Li, Meilian Li, Jiajun Nie, Ming Xu, Hao Feng, Liangsheng Xu, Cong Jiang, Qingmei Guan, Lili Huang, Valsa mali PR1-like protein modulates an apple valine-glutamine protein to suppress JA signaling-mediated immunity, 2024, 194, 0032-0889, 2755, 10.1093/plphys/kiae020 |


Figures(2)

Nick K. Olrichs, J. Bernd Helms. Novel insights into the function of the conserved domain of the CAP superfamily of proteins[J]. AIMS Biophysics, 2016, 3(2): 232-246. doi: 10.3934/biophy.2016.2.232






 DownLoad:
DownLoad: 