Citation: Nita R. Shah, Keni Vidilaseris, Henri Xhaard, Adrian Goldman. Integral membrane pyrophosphatases: a novel drug target for human pathogens?[J]. AIMS Biophysics, 2016, 3(1): 171-194. doi: 10.3934/biophy.2016.1.171

| [1] |
Cooperman BS, Baykov AA, Lahti R (1992) Evolutionary conservation of the active site of soluble inorganic pyrophosphatase. Trends Biochem Sci 17: 262–266. doi: 10.1016/0968-0004(92)90406-Y

|
| [2] | Lahti R (1983) Microbial inorganic pyrophosphatases. Microbiol Rev 47: 169–178. |
| [3] | Klemme JH (1976) Regulation of intracellular pyrophosphatase-activity and conservation of the phosphoanhydride-energy of inorganic pyrophosphate in microbial metabolism. Z Naturforsch C 31: 544–550. |
| [4] | Heinonen JK (2001) Biological role of inorganic pyrophosphate: Springer Science & Business Media. |
| [5] | Terkeltaub RA (2001) Inorganic pyrophosphate generation and disposition in pathophysiology. Am J Physiol Cell Physiol 281: C1–C11. |
| [6] |
Kajander T, Kellosalo J, Goldman A (2013) Inorganic pyrophosphatases: one substrate, three mechanisms. FEBS Lett 587: 1863–1869. doi: 10.1016/j.febslet.2013.05.003
|
| [7] | Chen J, Brevet A, Fromant M, et al. (1990) Pyrophosphatase is essential for growth of Escherichia coli. J Bacteriol 172: 5686–5689. |
| [8] | Lundin M, Baltscheffsky H, Ronne H (1991) Yeast PPA2 gene encodes a mitochondrial inorganic pyrophosphatase that is essential for mitochondrial function. J Biol Chem 266: 12168–12172. |
| [9] |
Kankare J, Salminen T, Lahti R, et al. (1996) Structure of Escherichia coli inorganic pyrophosphatase at 2.2 A resolution. Acta Crystallogr D 52: 551–563. doi: 10.1107/S0907444996000376
|
| [10] | Arutiunian E, Terzian S, Voronova A, et al. (1981) X-Ray diffraction study of inorganic pyrophosphatase from baker's yeast at the 3 angstroms resolution (Russian). DoklAkadNauk Sssr 258: 1481. |
| [11] |
Kellosalo J, Kajander T, Kogan K, et al. (2012) The structure and catalytic cycle of a sodium-pumping pyrophosphatase. Science 337: 473–476. doi: 10.1126/science.1222505
|
| [12] |
Lin SM, Tsai JY, Hsiao CD, et al. (2012) Crystal structure of a membrane-embedded H+-translocating pyrophosphatase. Nature 484: 399–403. doi: 10.1038/nature10963
|
| [13] |
Luoto HH, Baykov AA, Lahti R, et al. (2013) Membrane-integral pyrophosphatase subfamily capable of translocating both Na+ and H+. P Natl Acad Sci U S A 110: 1255–1260. doi: 10.1073/pnas.1217816110
|
| [14] |
Garcia-Contreras R, Celis H, Romero I (2004) Importance of Rhodospirillum rubrum H+-pyrophosphatase under low-energy conditions. J Bacteriol 186: 6651–6655. doi: 10.1128/JB.186.19.6651-6655.2004
|
| [15] |
Serrano A, Pérez-Castiñeira JR, Baltscheffsky M, et al. (2007) H+-PPases: yesterday, today and tomorrow. IUBMB Life 59: 76–83. doi: 10.1080/15216540701258132
|
| [16] |
Baykov AA, Malinen AM, Luoto HH, et al. (2013) Pyrophosphate-fueled Na+ and H+ transport in prokaryotes. Microbiol Mol Biol Rev 77: 267–276. doi: 10.1128/MMBR.00003-13
|
| [17] | Taiz L (1992) The Plant Vacuole. J Exp Biol 172: 113–122. |
| [18] | Maeshima M, Yoshida S (1989) Purification and properties of vacuolar membrane proton-translocating inorganic pyrophosphatase from mung bean. J Biol Chem 264: 20068–20073. |
| [19] | Belogurov GA, Lahti R (2002) A lysine substitute for K+. A460K mutation eliminates K+ dependence in H+-pyrophosphatase of Carboxydothermus hydrogenoformans. J Biol Chem 277: 49651–49654. |
| [20] |
Luoto HH, Belogurov GA, Baykov AA, et al. (2011) Na+-translocating Membrane Pyrophosphatases Are Widespread in the Microbial World and Evolutionarily Precede H+-translocating Pyrophosphatases. J Biol Chem 286: 21633–21642. doi: 10.1074/jbc.M111.244483
|
| [21] |
Luoto HH, Nordbo E, Malinen AM, et al. (2015) Evolutionarily divergent, Na+-regulated H+-transporting membrane-bound pyrophosphatases. Biochem J 467: 281–291. doi: 10.1042/BJ20141434
|
| [22] |
Tsai JY, Kellosalo J, Sun YJ, et al. (2014) Proton/sodium pumping pyrophosphatases: the last of the primary ion pumps. Curr Opin Struct Biol 27: 38–47. doi: 10.1016/j.sbi.2014.03.007
|
| [23] |
Drozdowicz YM, Rea PA (2001) Vacuolar H+ pyrophosphatases: from the evolutionary backwaters into the mainstream. Trends Plant Sci 6: 206–211. doi: 10.1016/S1360-1385(01)01923-9
|
| [24] | Björn LO (2015) The evolution of photosynthesis and its environmental impact. Photobiology: Springer. pp. 207–230. |
| [25] |
Kriegel A, Andres Z, Medzihradszky A, et al. (2015) Job Sharing in the Endomembrane System: Vacuolar Acidification Requires the Combined Activity of V-ATPase and V-PPase. Plant Cell 27: 3383–3396 doi: 10.1105/tpc.15.00733
|
| [26] |
Nakayasu T, Kawauchi, K., Hirata, H., et al. (1999) Cycloprodigiosin hydrochloride inhibits acidification of the plant vacuole. Plant Cell Physiol 40: 143–148. doi: 10.1093/oxfordjournals.pcp.a029521
|
| [27] | Bethmann B, Thaler M, Simonis W, et al. (1995) Electrochemical Potential Gradients of H+, K+, Ca2+, and Cl- across the Tonoplast of the Green Alga Eremosphaera Viridis. Plant Physiol 109: 1317–1326. |
| [28] |
Schumaker KS, Sze H (1990) Solubilization and reconstitution of the oat root vacuolar H+/Ca2+ exchanger. Plant Physiol 92: 340–345. doi: 10.1104/pp.92.2.340
|
| [29] |
Jiang L, Phillips TE, Hamm CA, et al. (2001) The protein storage vacuole: a unique compound organelle. J Cell Biol 155: 991–1002. doi: 10.1083/jcb.200107012
|
| [30] |
Li J, Yang H, Peer WA, et al. (2005) Arabidopsis H+-PPase AVP1 regulates auxin-mediated organ development. Science 310: 121–125. doi: 10.1126/science.1115711
|
| [31] |
Sabatini S, Beis D, Wolkenfelt H, et al. (1999) An auxin-dependent distal organizer of pattern and polarity in the Arabidopsis root. Cell 99: 463–472. doi: 10.1016/S0092-8674(00)81535-4
|
| [32] | Brini F, Hanin M, Mezghani I, et al. (2007) Overexpression of wheat Na+/H+ antiporter TNHX1 and H+-pyrophosphatase TVP1 improve salt- and drought-stress tolerance in Arabidopsis thaliana plants. J Exp Bot 58: 301–308. |
| [33] |
Lv SL, Lian LJ, Tao PL, et al. (2009) Overexpression of Thellungiella halophila H+-PPase (TsVP) in cotton enhances drought stress resistance of plants. Planta 229: 899–910. doi: 10.1007/s00425-008-0880-4
|
| [34] | Li X, Guo C, Gu J, et al. (2014) Overexpression of VP, a vacuolar H+-pyrophosphatase gene in wheat (Triticum aestivum L.), improves tobacco plant growth under Pi and N deprivation, high salinity, and drought. J Exp Bot 65: 683–696. |
| [35] |
Park S, Li J, Pittman JK, et al. (2005) Up-regulation of a H+-pyrophosphatase (H+-PPase) as a strategy to engineer drought-resistant crop plants. P Natl Acad Sci U S A 102: 18830–18835. doi: 10.1073/pnas.0509512102
|
| [36] |
Battisti DS, Naylor RL (2009) Historical warnings of future food insecurity with unprecedented seasonal heat. Science 323: 240–244. doi: 10.1126/science.1164363
|
| [37] |
Zhang H, Shen G, Kuppu S, et al. (2011) Creating drought- and salt-tolerant cotton by overexpressing a vacuolar pyrophosphatase gene. Plant Signal Behav 6: 861–863. doi: 10.4161/psb.6.6.15223
|
| [38] | Yaeger R (1996) Protozoa: structure, classification, growth, and development. |
| [39] |
Kappe SH, Buscaglia CA, Bergman LW, et al. (2004) Apicomplexan gliding motility and host cell invasion: overhauling the motor model. Trends Parasitol 20: 13–16. doi: 10.1016/j.pt.2003.10.011
|
| [40] |
Murray CJL, Vos T, Lozano R, et al. (2012) Disability-adjusted life years (DALYs) for 291 diseases and injuries in 21 regions, 1990-2010: a systematic analysis for the Global Burden of Disease Study 2010. Lancet 380: 2197–2223. doi: 10.1016/S0140-6736(12)61689-4
|
| [41] | World Health Organization (2013) WHO Global malaria report 2013. Geneva, Switzerland: World Health Organization. |
| [42] |
Greenwood BM, Fidock DA, Kyle DE, et al. (2008) Malaria: progress, perils, and prospects for eradication. J Clin Invest 118: 1266–1276. doi: 10.1172/JCI33996
|
| [43] |
Crompton PD, Moebius J, Portugal S, et al. (2014) Malaria immunity in man and mosquito: insights into unsolved mysteries of a deadly infectious disease. Annu Rev Immunol 32: 157–187. doi: 10.1146/annurev-immunol-032713-120220
|
| [44] |
Smith T, Schellenberg JA, Hayes R (1994) Attributable fraction estimates and case definitions for malaria in endemic areas. Stat Med 13: 2345–2358. doi: 10.1002/sim.4780132206
|
| [45] | World Health Organization (2014) WHO global malaria report 2014. Geneva, Switzerland: World Health Organization. |
| [46] |
Shortt HE, Fairley NH, Covell G, et al. (1951) The pre-erythrocytic stage of Plasmodium falciparum. Trans R Soc Trop Med Hyg 44: 405–419. doi: 10.1016/S0035-9203(51)80019-1
|
| [47] |
Mota MM, Pradel G, Vanderberg JP, et al. (2001) Migration of Plasmodium sporozoites through cells before infection. Science 291: 141–144. doi: 10.1126/science.291.5501.141
|
| [48] |
Sturm A, Amino R, van de Sand C, et al. (2006) Manipulation of host hepatocytes by the malaria parasite for delivery into liver sinusoids. Science 313: 1287–1290. doi: 10.1126/science.1129720
|
| [49] | Kollien A, Meyer H, Großpietsch T, et al. (1998) Correlation of the development of Trypanosoma cruzi with the physiology in the rectum of the vector. Parasitol Int 47: 143. |
| [50] |
Kollien A, Schaub G (2000) The development of Trypanosoma cruzi in triatominae. Parasitol Today 16: 381–387. doi: 10.1016/S0169-4758(00)01724-5
|
| [51] |
Docampo R, Jimenez V, Lander N, et al. (2013) New insights into roles of acidocalcisomes and contractile vacuole complex in osmoregulation in protists. Int Rev Cell Mol Biol 305: 69–113. doi: 10.1016/B978-0-12-407695-2.00002-0
|
| [52] |
Docampo R, Scott DA, Vercesi AE, et al. (1995) Intracellular Ca2+ storage in acidocalcisomes of Trypanosoma cruzi. Biochem J 310: 1005–1012. doi: 10.1042/bj3101005
|
| [53] |
Garcia CR, Ann SE, Tavares ES, et al. (1998) Acidic calcium pools in intraerythrocytic malaria parasites. Eur J Cell Biol 76: 133–138. doi: 10.1016/S0171-9335(98)80026-5
|
| [54] |
Scott DA, Moreno S, Docampo R (1995) Ca2+ storage in Trypanosoma brucei: the influence of cytoplasmic pH and importance of vacuolar acidity. Biochem J 310: 789–794. doi: 10.1042/bj3100789
|
| [55] |
Moreno S, Zhong L (1996) Acidocalcisomes in Toxoplasma gondii tachyzoites. Biochem J 313: 655–659. doi: 10.1042/bj3130655
|
| [56] |
Ruiz FA, Rodrigues CO, Docampo R (2001) Rapid Changes in Polyphosphate Content within Acidocalcisomes in Response to Cell Growth, Differentiation, and Environmental Stress in Trypanosoma cruzi. J Biol Chem 276: 26114–26121. doi: 10.1074/jbc.M102402200
|
| [57] |
Ruiz FA, Marchesini N, Seufferheld M, et al. (2001) The polyphosphate bodies of Chlamydomonas reinhardtii possess a proton-pumping pyrophosphatase and are similar to acidocalcisomes. J Biol Chem 276: 46196–46203. doi: 10.1074/jbc.M105268200
|
| [58] |
Lemercier G, Dutoya S, Luo S, et al. (2002) A vacuolar-type H+-pyrophosphatase governs maintenance of functional acidocalcisomes and growth of the insect and mammalian forms of Trypanosoma brucei. J Biol Chem 277: 37369–37376. doi: 10.1074/jbc.M204744200
|
| [59] |
Vercesi AE, Moreno S, Docampo R (1994) Ca2+/H+ exchange in acidic vacuoles of Trypanosoma brucei. Biochem J 304: 227–233. doi: 10.1042/bj3040227
|
| [60] |
Rohloff P, Montalvetti A, Docampo R (2004) Acidocalcisomes and the contractile vacuole complex are involved in osmoregulation in Trypanosoma cruzi. J Biol Chem 279: 52270–52281. doi: 10.1074/jbc.M410372200
|
| [61] |
Scott DA, de Souza W, Benchimol M, et al. (1998) Presence of a plant-like proton-pumping pyrophosphatase in acidocalcisomes of Trypanosoma cruzi. J Biol Chem 273: 22151–22158. doi: 10.1074/jbc.273.34.22151
|
| [62] |
Marchesini N, Luo S, Rodrigues C, et al. (2000) Acidocalcisomes and a vacuolar H+-pyrophosphatase in malaria parasites. Biochem J 347: 243–253. doi: 10.1042/bj3470243
|
| [63] |
Lu H-G, Zhong L, de Souza W, et al. (1998) Ca2+ content and expression of an acidocalcisomal calcium pump are elevated in intracellular forms of Trypanosoma cruzi. Mol Cell Biol 18: 2309–2323. doi: 10.1128/MCB.18.4.2309
|
| [64] |
Liu J, Pace D, Dou Z, et al. (2014) A vacuolar H+ pyrophosphatase (TgVP1) is required for microneme secretion, host cell invasion, and extracellular survival of Toxoplasma gondii. Molecular microbiology 93: 698–712. doi: 10.1111/mmi.12685
|
| [65] | Rodrigues CO, Scott DA, Bailey BN, et al. (2000) Vacuolar proton pyrophosphatase activity and pyrophosphate (PPi) in Toxoplasma gondii as possible chemotherapeutic targets. Biochem J 349 Pt 3: 737–745. |
| [66] | Martin MB, Grimley JS, Lewis JC, et al. (2001) Bisphosphonates Inhibit the Growth of Trypanosoma brucei. Trypanosoma cruzi. Leishmania donovani. Toxoplasma gondii, and Plasmodium falciparum: A Potential Route to Chemotherapy. J Med Chem 44: 909–916. |
| [67] |
Martin MB, Sanders JM, Kendrick H, et al. (2002) Activity of bisphosphonates against Trypanosoma brucei rhodesiense. J Med Chem 45: 2904–2914. doi: 10.1021/jm0102809
|
| [68] |
Rodan GA, Martin TJ (2000) Therapeutic approaches to bone diseases. Science 289: 1508–1514. doi: 10.1126/science.289.5484.1508
|
| [69] |
Baykov AA, Dubnova EB, Bakuleva NP, et al. (1993) Differential sensitivity of membrane-associated pyrophosphatases to inhibition by diphosphonates and fluoride delineates two classes of enzyme. FEBS Lett 327: 199–202. doi: 10.1016/0014-5793(93)80169-U
|
| [70] |
Smirnova IN, Kudryavtseva NA, Komissarenko SV, et al. (1988) Diphosphonates are potent inhibitors of mammalian inorganic pyrophosphatase. Arch Biochem Biophys 267: 280–284. doi: 10.1016/0003-9861(88)90033-1
|
| [71] | Holdeman L, Good I, Moore W (1976) Human fecal flora: variation in bacterial composition within individuals and a possible effect of emotional stress. Appl Environ Microbiol 31: 359–375. |
| [72] |
Salyers A (1984) Bacteroides of the human lower intestinal tract. Annu Rev Microbiol 38: 293–313. doi: 10.1146/annurev.mi.38.100184.001453
|
| [73] |
Xu J, Gordon JI (2003) Honor thy symbionts. P Natl Acad Sci USA 100: 10452–10459. doi: 10.1073/pnas.1734063100
|
| [74] |
Bäckhed F, Ley RE, Sonnenburg JL, et al. (2005) Host-bacterial mutualism in the human intestine. Science 307: 1915–1920. doi: 10.1126/science.1104816
|
| [75] |
Brook I, Johnson N, Overturf GD, et al. (1977) Mixed bacterial meningitis: A complication of ventriculo-and lumboperitoneal shunts: Report of two cases. J Neurosurg 47: 961–964. doi: 10.3171/jns.1977.47.6.0961
|
| [76] | Odugbemi T, Jatto S, Afolabi K (1985) Bacteroides fragilis meningitis. J Clin Microbiol 21: 282–283. |
| [77] |
Wexler HM (2007) Bacteroides: the good, the bad, and the nitty-gritty. Clin Microbiol Rev 20: 593-621. doi: 10.1128/CMR.00008-07
|
| [78] |
Goldstein EJ (1996) Anaerobic bacteremia. Clin Infect Dis 23: S97–S101. doi: 10.1093/clinids/23.Supplement_1.S97
|
| [79] |
Hecht DW (2004) Prevalence of antibiotic resistance in anaerobic bacteria: worrisome developments. Clin Infect Dis 39: 92–97. doi: 10.1086/421558
|
| [80] |
Nguyen MH, Victor LY, Morris AJ, et al. (2000) Antimicrobial resistance and clinical outcome of Bacteroides bacteremia: findings of a multicenter prospective observational trial. Clin Infect Dis 30: 870–876. doi: 10.1086/313805
|
| [81] |
Yoon H-S, Kim S-Y, Kim I-S (2013) Stress response of plant H+-PPase-expressing transgenic Escherichia coli and Saccharomyces cerevisiae: a potentially useful mechanism for the development of stress-tolerant organisms. J Appl Genet 54: 129–133. doi: 10.1007/s13353-012-0117-x
|
| [82] | Rafii F, Sutherland JB, Cerniglia CE (2008) Effects of treatment with antimicrobial agents on the human colonic microflora. Ther Clin Risk Manag 4: 1343. |
| [83] |
Owens RC, Donskey CJ, Gaynes RP, et al. (2008) Antimicrobial-associated risk factors for Clostridium difficile infection. Clin Infect Dis 46: S19–S31. doi: 10.1086/521859
|
| [84] |
Hopkins M, Macfarlane G (2002) Changes in predominant bacterial populations in human faeces with age and with Clostridium difficile infection. J Med Microbiol 51: 448–454. doi: 10.1099/0022-1317-51-5-448
|
| [85] |
Gaxiola RA, Palmgren MG, Schumacher K (2007) Plant proton pumps. FEBS Lett 581: 2204–2214. doi: 10.1016/j.febslet.2007.03.050
|
| [86] |
Maeshima M (2000) Vacuolar H+-pyrophosphatase. Biochim Biophys Acta 1465: 37–51. doi: 10.1016/S0005-2736(00)00130-9
|
| [87] |
Asaoka M, Segami S, Maeshima M (2014) Identification of the critical residues for the function of vacuolar H+-pyrophosphatase by mutational analysis based on the 3D structure. J Biochem 156: 333–344. doi: 10.1093/jb/mvu046
|
| [88] | Terstappen GC, Reggiani A (2001) In silico research in drug discovery. Trends Pharmacol Sci 22: 23–26. |
| [89] |
Rask-Andersen M, Almen MS, Schioth HB (2011) Trends in the exploitation of novel drug targets. Nat Rev Drug Discov 10: 579–590. doi: 10.1038/nrd3478
|
| [90] |
Wishart DS, Knox C, Guo AC, et al. (2006) DrugBank: a comprehensive resource for in silico drug discovery and exploration. Nucleic Acids Res 34: D668–672. doi: 10.1093/nar/gkj067
|
| [91] |
Lipinski CA, Lombardo F, Dominy BW, et al. (1997) Experimental and computational approaches to estimate solubility and permeability in drug discovery and development settings. Adv Drug Deliver Rev 23: 3–25. doi: 10.1016/S0169-409X(96)00423-1
|
| [92] | Biologic E, Foa P, Zak B (1967) Microdetermination of inorganic phosphate, phospholipids, and total phosphate in biologic materials. Clin Chem 13: 326–332. |
| [93] |
Hajduk PJ, Huth JR, Fesik SW (2005) Druggability indices for protein targets derived from NMR-based screening data. J Med Chem 48: 2518–2525. doi: 10.1021/jm049131r
|
| [94] |
Hajduk PJ, Huth JR, Tse C (2005) Predicting protein druggability. Drug Discov Today 10: 1675–1682. doi: 10.1016/S1359-6446(05)03624-X
|
| [95] |
Shuker SB, Hajduk PJ, Meadows RP, et al. (1996) Discovering high-affinity ligands for proteins: SAR by NMR. Science 274: 1531–1534. doi: 10.1126/science.274.5292.1531
|
| [96] |
Andrews SP, Brown GA, Christopher JA (2014) Structure-Based and Fragment-Based GPCR Drug Discovery. ChemMedChem 9: 256–275. doi: 10.1002/cmdc.201300382
|
| [97] |
Congreve M, Rich RL, Myszka DG, et al. (2011) 5 Fragment Screening of Stabilized G-Protein-Coupled Receptors Using Biophysical Methods. Methods Enzymol 493: 115. doi: 10.1016/B978-0-12-381274-2.00005-4
|
| [98] | Hawkins PCD (2006) A comparison of structure-based and shape-based tools for virtual screening. Abstr Pap Am Chem S 231. |
| [99] |
Lyne PD (2002) Structure-based virtual screening: an overview. Drug Discov Today 7: 1047–1055. doi: 10.1016/S1359-6446(02)02483-2
|
| [100] |
Huang N, Shoichet BK, Irwin JJ (2006) Benchmarking sets for molecular docking. J Med Chem 49: 6789–6801. doi: 10.1021/jm0608356
|
| [101] |
Aqvist J, Luzhkov VB, Brandsdal BO (2002) Ligand binding affinities from MD simulations. Acc Chem Res 35: 358–365. doi: 10.1021/ar010014p
|
| [102] |
Bohm HJ (1998) Prediction of binding constants of protein ligands: A fast method for the prioritization of hits obtained from de novo design or 3D database search programs. J Comput Aid Mol Des 12: 309–323. doi: 10.1023/A:1007999920146
|
| [103] |
Muegge I, Martin YC (1999) A general and fast scoring function for protein-ligand interactions: A simplified potential approach. J Med Chem 42: 791–804. doi: 10.1021/jm980536j
|
| [104] |
Kuntz ID, Blaney JM, Oatley SJ, et al. (1982) A geometric approach to macromolecule-ligand interactions. J Mol Biol 161: 269–288. doi: 10.1016/0022-2836(82)90153-X
|
| [105] | Morris GM, Goodsell DS, Halliday RS, et al. (1998) Automated docking using a Lamarckian genetic algorithm and an empirical binding free energy function. J Comput Chem 19: 1639–1662. |
| [106] |
Verdonk ML, Cole JC, Hartshorn MJ, et al. (2003) Improved protein-ligand docking using GOLD. Proteins 52: 609–623. doi: 10.1002/prot.10465
|
| [107] |
Bohm HJ (1992) The computer program LUDI: a new method for the de novo design of enzyme inhibitors. J Comput Aid Mol Des 6: 61–78. doi: 10.1007/BF00124387
|
| [108] |
Korb O, Stutzle T, Exner TE (2009) Empirical scoring functions for advanced protein-ligand docking with PLANTS. J Chem Inf Model 49: 84–96. doi: 10.1021/ci800298z
|
| [109] |
Ishchenko AV, Shakhnovich EI (2002) SMall molecule growth 2001 (SMoG2001): An improved knowledge-based scoring function for protein-ligand interactions. J Med Chem 45: 2770–2780. doi: 10.1021/jm0105833
|
| [110] | Triballeau N, Acher F, Brabet I, et al. (2005) Virtual screening workflow development guided by the “receiver operating characteristic” curve approach. Application to high-throughput docking on metabotropic glutamate receptor subtype 4. J of M Chem 48: 2534–2547. |
| [111] |
Schneider G, Fechner U (2005) Computer-based de novo design of drug-like molecules. Nat Rev Drug Discov 4: 649–663. doi: 10.1038/nrd1799
|
| [112] |
Miranker A, Karplus M (1991) Functionality maps of binding sites: a multiple copy simultaneous search method. Proteins 11: 29–34. doi: 10.1002/prot.340110104
|
| [113] |
Wang R, Gao Y, Lai L (2000) LigBuilder: A Multi-Purpose Program for Structure-Based Drug Design. J Mol Model 6: 498–516. doi: 10.1007/s0089400060498
|
| [114] |
Gillet V, Johnson AP, Mata P, et al. (1993) SPROUT: a program for structure generation. J Comput Aid Mol Des 7: 127–153. doi: 10.1007/BF00126441
|
| [115] |
Lolicato M, Bucchi A, Arrigoni C, et al. (2014) Cyclic dinucleotides bind the C-linker of HCN4 to control channel cAMP responsiveness. Nat Chemical Biol 10: 457–462. doi: 10.1038/nchembio.1521
|
| [116] |
Elliott TS, Slowey A, Ye Y, et al. (2012) The use of phosphate bioisosteres in medicinal chemistry and chemical biology. MedChemComm 3: 735–751. doi: 10.1039/c2md20079a
|
| [117] |
Rye C, Baell J (2005) Phosphate isosteres in medicinal chemistry. Curr Med Chem 12: 3127–3141. doi: 10.2174/092986705774933452
|
| [118] |
Manly CJ, Chandrasekhar J, Ochterski JW, et al. (2008) Strategies and tactics for optimizing the Hit-to-Lead process and beyond—A computational chemistry perspective. Drug Discov Today 13: 99–109. doi: 10.1016/j.drudis.2007.10.019
|
| [119] | Garbaccio RM, Parmee ER (2016) The Impact of Chemical Probes in Drug Discovery: A Pharmaceutical Industry Perspective. Chem Biol 23: 10–17. |
| [120] |
Zhang Y, Zhu W, Liu Y-L, et al. (2013) Chemo-immunotherapeutic antimalarials targeting isoprenoid biosynthesis. ACS Med Chem Lett 4: 423–427. doi: 10.1021/ml4000436
|
| [121] |
Jahnke W, Rondeau J-M, Cotesta S, et al. (2010) Allosteric non-bisphosphonate FPPS inhibitors identified by fragment-based discovery. Nat Chem Biol 6: 660–666. doi: 10.1038/nchembio.421
|
| [122] |
Liu Y-L, Lindert S, Zhu W, et al. (2014) Taxodione and arenarone inhibit farnesyl diphosphate synthase by binding to the isopentenyl diphosphate site. P Nat Acad Sci U S A 111: E2530–E2539. doi: 10.1073/pnas.1409061111
|
| [123] |
Lindert S, Zhu W, Liu Y-L, et al. (2013) Farnesyl diphosphate synthase inhibitors from in silico screening. Chem Biol Drug Des 81: 742–748. doi: 10.1111/cbdd.12121
|
| [124] |
Itaya K, Ui M (1966) A new micromethod for the colorimetric determination of inorganic phosphate. Clin Chim Acta 14: 361–366. doi: 10.1016/0009-8981(66)90114-8
|
| [125] |
Llona-Minguez S, Höglund A, Jacques S, et al. (2016) Discovery of the first potent and selective inhibitors of the human dCTP pyrophosphatase 1 (dCTPase). J Med Chem 59: 1140–1148. doi: 10.1021/acs.jmedchem.5b01741
|
| [126] | Zhen RG, Baykov AA, Bakuleva NP, et al. (1994) Aminomethylenediphosphonate: A Potent Type-Specific Inhibitor of Both Plant and Phototrophic Bacterial H+-Pyrophosphatases. Plant Physiol 104: 153–159. |
| [127] |
Heikinheimo P, Tuominen V, Ahonen AK, et al. (2001) Toward a quantum-mechanical description of metal-assisted phosphoryl transfer in pyrophosphatase. P Natl Acad Sci U S A 98: 3121–3126. doi: 10.1073/pnas.061612498
|
| [128] |
Falagas ME, Kasiakou SK (2006) Toxicity of polymyxins: a systematic review of the evidence from old and recent studies. Crit Care 10: R27. doi: 10.1186/cc3995
|
| [129] |
Avent M, Rogers B, Cheng A, et al. (2011) Current use of aminoglycosides: indications, pharmacokinetics and monitoring for toxicity. Intern Med J 41: 441–449. doi: 10.1111/j.1445-5994.2011.02452.x
|
| [130] |
Tseng YY, Dupree C, Chen ZJ, et al. (2009) SplitPocket: identification of protein functional surfaces and characterization of their spatial patterns. Nucleic Acids Res 37: W384–W389. doi: 10.1093/nar/gkp308
|
| [131] |
Tseng YY, Li WH (2009) Identification of protein functional surfaces by the concept of a split pocket. Proteins 76: 959–976. doi: 10.1002/prot.22402
|
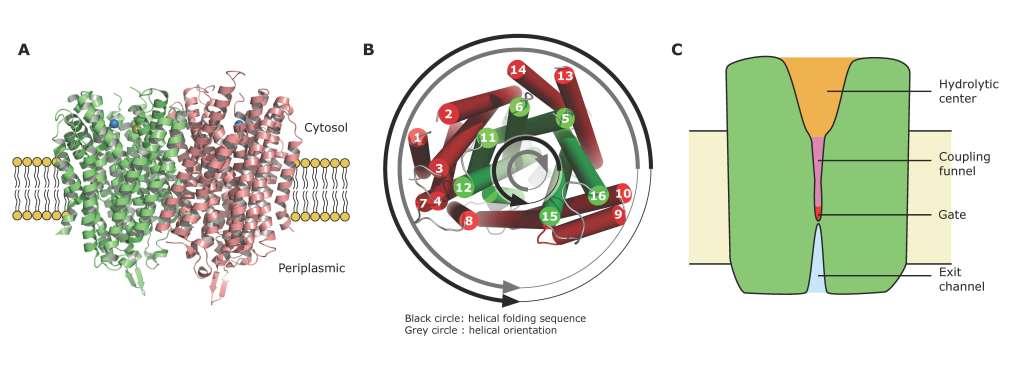
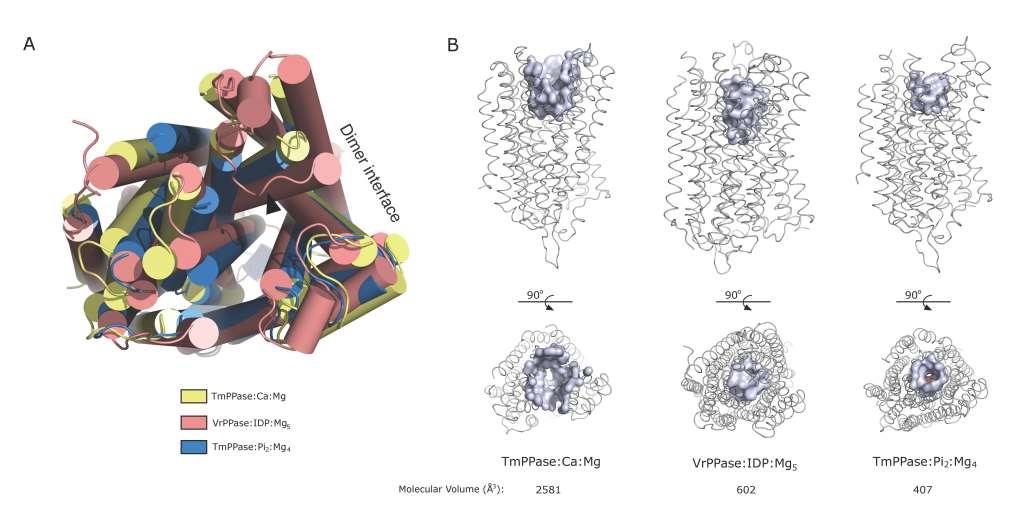
Figures(2) / Tables(1)

Nita R. Shah, Keni Vidilaseris, Henri Xhaard, Adrian Goldman. Integral membrane pyrophosphatases: a novel drug target for human pathogens?[J]. AIMS Biophysics, 2016, 3(1): 171-194. doi: 10.3934/biophy.2016.1.171







 DownLoad:
DownLoad: