Citation: Shen Helvig, Intan D. M. Azmi, Seyed M. Moghimi, Anan Yaghmur. Recent Advances in Cryo-TEM Imaging of Soft Lipid Nanoparticles[J]. AIMS Biophysics, 2015, 2(2): 116-130. doi: 10.3934/biophy.2015.2.116

| [1] |
Moghimi SM, Hunter AC, Murray JC (2005) Nanomedicine: current status and future prospects. FASEB J 19: 311-330. doi: 10.1096/fj.04-2747rev

|
| [2] |
Grazú V, Moros M, Sánchez-Espinel C (2012) Nanobiotechnology - Inorganic Nanoparticles vs Organic Nanoparticles. Front Nanosci 4: 337-440. doi: 10.1016/B978-0-12-415769-9.00014-5
|
| [3] |
Lim SB, Banerjee A, Önyüksel H (2012) Improvement of drug safety by the use of lipid-based nanocarriers. J Control Release 163: 34-45. doi: 10.1016/j.jconrel.2012.06.002
|
| [4] |
Nel AE, Maedler L, Velegol D, et al. (2009) Understanding biophysicochemical interactions at the nano-bio interface. Nat Mater 8: 543-557. doi: 10.1038/nmat2442
|
| [5] |
Couvreur P, Vauthier C (2006) Nanotechnology: intelligent design to treat complex disease. Pharm Res 23: 1417-1450. doi: 10.1007/s11095-006-0284-8
|
| [6] | Jin S-E, Jin H-E, Hong S-S (2014) Targeted delivery system of nanobiomaterials in anticancer therapy: from cells to clinics. BioMed Res Int 2014: 814208. |
| [7] |
Moghimi SM, Peer D, Langer R (2011) Reshaping the future of nanopharmaceuticals: adiudicium. ACS Nano 5: 8454-8458. doi: 10.1021/nn2038252
|
| [8] |
Klang V, Valenta C, Matsko NB (2013) Electron microscopy of pharmaceutical systems. Micron 44: 45-74. doi: 10.1016/j.micron.2012.07.008
|
| [9] |
Danino D (2012) Cryo-TEM of soft molecular assemblies. Curr Opin Colloid Interface Sci 17: 316-329. doi: 10.1016/j.cocis.2012.10.003
|
| [10] | Danino D (2001) Digital cryogenic transmission electron microscopy: an advanced tool for direct imaging of complex fluids. Colloids Surf A 183-185: 113-122. |
| [11] |
Burrows ND, Penn RL (2013) Cryogenic Transmission Electron Microscopy: Aqueous Suspensions of Nanoscale Objects. Microsc Microanal 19: 1542-1553. doi: 10.1017/S1431927613013354
|
| [12] |
Cho EJ, Holback H, Liu KC, et al. (2013) Nanoparticle characterization: State of the art, challenges, and emerging technologies. Mol Pharm 10: 2093-2110. doi: 10.1021/mp300697h
|
| [13] |
McNeil SE (2011) Challenges for nanoparticle characterization. Methods Mol Biol 697: 9-15. doi: 10.1007/978-1-60327-198-1_2
|
| [14] |
Friedrich H, Frederik PM, de With G, et al. (2010) Imaging of self-assembled structures: interpretation of TEM and cryo-TEM images. Angew Chem Int Ed Engl 49: 7850-7858. doi: 10.1002/anie.201001493
|
| [15] |
Kuntsche J, Horst JC, Bunjes H (2011) Cryogenic transmission electron microscopy (cryo-TEM) for studying the morphology of colloidal drug delivery systems. Int J Pharm 417: 120-137. doi: 10.1016/j.ijpharm.2011.02.001
|
| [16] |
Rubino S, Akhtar S, Melin P, et al. (2012) A site-specific focused-ion-beam lift-out method for cryo Transmission Electron Microscopy. J Struct Biol 180: 572-576. doi: 10.1016/j.jsb.2012.08.012
|
| [17] | Frederik PM, Stuart MC, Bomans PH, et al. (1991) Perspective and limitations of cryo-electron microscopy. From model systems to biological specimens. J Microsc 161: 253-262. |
| [18] | Mielanczyk L, Matysiak N, Michalski M, et al. (2014) Closer to the native state. Critical evaluation of cryo-techniques for Transmission Electron Microscopy: preparation of biological samples. Folia Histochem Cytobiol 52: 1-17. |
| [19] |
Pilhofer M, Ladinsky MS, McDowall AW, et al. (2010) Bacterial TEM: new insights from cryo-microscopy. Methods Cell Biol 96: 21-45. doi: 10.1016/S0091-679X(10)96002-0
|
| [20] |
De Carlo S, Harris JR (2011) Negative staining and cryo-negative staining of macromolecules and viruses for TEM. Micron 42: 117-131. doi: 10.1016/j.micron.2010.06.003
|
| [21] |
Adrian M, Dubochet J, Lepault J, et al. (1984) Cryo-electron microscopy of viruses. Nature 308: 32-36. doi: 10.1038/308032a0
|
| [22] | Fatouros DG, Müllertz A (2013) Development of Self-Emulsifying Drug Delivery Systems (SEDDS) for Oral Bioavailability Enhancement of Poorly Soluble Drugs. Drug Delivery Strategies for Poorly Water-Soluble Drugs: John Wiley & Sons Ltd. pp. 225-245. |
| [23] | Talmon Y (2007) Seeing giant micelles by cryogenic-temperature transmission electron microscopy (cryo-TEM). Surfactant Sci Ser 140: 163. |
| [24] |
Spernath L, Regev O, Levi-Kalisman Y, et al. (2009) Phase transitions in O/W lauryl acrylate emulsions during phase inversion, studied by light microscopy and cryo-TEM. Colloids Surf A 332: 19-25. doi: 10.1016/j.colsurfa.2008.08.026
|
| [25] |
Almgren M, Edwards K, Karlsson G (2000) Cryo transmission electron microscopy of liposomes and related structures. Colloids Surf A 174: 3-21. doi: 10.1016/S0927-7757(00)00516-1
|
| [26] |
Ferreira DA, Bentley MVLB, Karlsson G, et al. (2006) Cryo-TEM investigation of phase behaviour and aggregate structure in dilute dispersions of monoolein and oleic acid. Int J Pharm 310: 203-212. doi: 10.1016/j.ijpharm.2005.11.028
|
| [27] |
Wytrwal M, Bednar J, Nowakowska M, et al. (2014) Interactions of serum with polyelectrolyte-stabilized liposomes: Cryo-TEM studies. Colloids Surf B 120: 152-159. doi: 10.1016/j.colsurfb.2014.02.040
|
| [28] |
Fatouros DG, Bergenstahl B, Mullertz A (2007) Morphological observations on a lipid-based drug delivery system during in vitro digestion. Eur J Pharm Sci 31: 85-94. doi: 10.1016/j.ejps.2007.02.009
|
| [29] |
Basáñez G, Ruiz-Argüello MB, Alonso A, et al. (1997) Morphological changes induced by phospholipase C and by sphingomyelinase on large unilamellar vesicles: a cryo-transmission electron microscopy study of liposome fusion. Biophys J 72: 2630-2637. doi: 10.1016/S0006-3495(97)78906-9
|
| [30] |
Phan S, Hawley A, Mulet X, et al. (2013) Structural aspects of digestion of medium chain triglycerides studied in real time using sSAXS and Cryo-TEM. Pharm Res 30: 3088-3100. doi: 10.1007/s11095-013-1108-2
|
| [31] |
Alfredsson V (2005) Cryo-TEM studies of DNA and DNA-lipid structures. Curr Opin Colloid Interface Sci 10: 269-273. doi: 10.1016/j.cocis.2005.09.005
|
| [32] |
Abraham SA, Edwards K, Karlsson G, et al. (2004) An evaluation of transmembrane ion gradient-mediated encapsulation of topotecan within liposomes. J Control Release 96: 449-461. doi: 10.1016/j.jconrel.2004.02.017
|
| [33] |
Johansson E, Lundquist A, Zuo S, et al. (2007) Nanosized bilayer disks: attractive model membranes for drug partition studies. Biochim Biophys Acta 1768: 1518-1525. doi: 10.1016/j.bbamem.2007.03.006
|
| [34] | Yaghmur A, Glatter O (2009) Characterization and potential applications of nanostructured aqueous dispersions. Adv Colloid Interface Sci 147-148: 333-342. |
| [35] | Yaghmur A, Glatter O (2012) Self-Assembly in Lipidic Particles. Self-Assembled Supramolecular Architectures: Lyotropic Liquid Crystals: John Wiley & Sons pp. 129-155. |
| [36] |
Yaghmur A, Rappolt M (2012) Structural characterization of lipidic systems under nonequilibrium conditions. Eur Biophys J 41: 831-840. doi: 10.1007/s00249-012-0815-7
|
| [37] |
Nilsson C, Edwards K, Eriksson J, et al. (2012) Characterization of oil-free and oil-loaded liquid-crystalline particles stabilized by negatively charged stabilizer citrem. Langmuir 28: 11755-11766. doi: 10.1021/la3021244
|
| [38] |
Hedegaard SF, Nilsson C, Laurinmaki P, et al. (2013) Nanostructured aqueous dispersions of citrem interacting with lipids and PEGylated lipids. RSC Adv 3: 24576-24585. doi: 10.1039/c3ra44583f
|
| [39] |
Chang DP, Jankunec M, Barauskas J, et al. (2012) Adsorption of lipid liquid crystalline nanoparticles: effects of particle composition, internal structure, and phase behavior. Langmuir 28: 10688-10696. doi: 10.1021/la301579g
|
| [40] |
Siegel DP, Burns JL, Chestnut MH, et al. (1989) Intermediates in membrane fusion and bilayer/nonbilayer phase transitions imaged by time-resolved cryo-transmission electron microscopy. Biophys J 56: 161-169. doi: 10.1016/S0006-3495(89)82661-X
|
| [41] |
Barauskas J, Johnsson M, Tiberg F (2005) Self-assembled lipid superstructures: beyond vesicles and liposomes. Nano Lett 5: 1615-1619. doi: 10.1021/nl050678i
|
| [42] |
Janiak J, Bayati S, Galantini L, et al. (2012) Nanoparticles with a bicontinuous cubic internal structure formed by cationic and non-ionic surfactants and an anionic polyelectrolyte. Langmuir 28: 16536-16546. doi: 10.1021/la303938k
|
| [43] |
Yaghmur A, De Campo L, Sagalowicz L, et al. (2005) Emulsified microemulsions and oil-containing liquid crystalline phases. Langmuir 21: 569-577. doi: 10.1021/la0482711
|
| [44] |
Yaghmur A, de Campo L, Salentinig S, et al. (2006) Oil-loaded monolinolein-based particles with confined inverse discontinuous cubic structure (Fd3m). Langmuir 22: 517-521. doi: 10.1021/la052109w
|
| [45] |
de Campo L, Yaghmur A, Sagalowicz L, et al. (2004) Reversible Phase Transitions in Emulsified Nanostructured Lipid Systems. Langmuir 20: 5254-5261. doi: 10.1021/la0499416
|
| [46] |
Nilsson C, Østergaard J, Larsen SW, et al. (2014) PEGylation of phytantriol-based lyotropic liquid crystalline particles-the effect of lipid composition, PEG chain length, and temperature on the internal nanostructure. Langmuir 30: 6398-6407. doi: 10.1021/la501411w
|
| [47] |
Sagalowicz L, Michel M, Adrian M, et al. (2006) Crystallography of dispersed liquid crystalline phases studied by cryo-transmission electron microscopy. J Microsc 221: 110-121. doi: 10.1111/j.1365-2818.2006.01544.x
|
| [48] |
Gao M, Kim Y-K, Zhang C, et al. (2014) Direct observation of liquid crystals using cryo-TEM: specimen preparation and low-dose imaging. Microsc Res Tech 77: 754-772. doi: 10.1002/jemt.22397
|
| [49] |
Angelova A, Angelov B, Drechsler M, et al. (2013) Protein entrapment in PEGylated lipid nanoparticles. Int J Pharm 454: 625-632. doi: 10.1016/j.ijpharm.2013.06.006
|
| [50] |
Sagalowicz L, Mezzenga R, Leser ME (2006) Investigating reversed liquid crystalline mesophases. Curr Opin Colloid Interface Sci 11: 224-229. doi: 10.1016/j.cocis.2006.07.002
|
| [51] |
Driever CD, Mulet X, Waddington LJ, et al. (2013) Layer-by-layer polymer coating on discrete particles of cubic lyotropic liquid crystalline dispersions (cubosomes). Langmuir 29: 12891-12900. doi: 10.1021/la401660h
|
| [52] |
Oliveira IMSC, Silva JPN, Feitosa E, et al. (2012) Aggregation behavior of aqueous dioctadecyldimethylammonium bromide/monoolein mixtures: a multitechnique investigation on the influence of composition and temperature. J Colloid Interface Sci 374: 206-217. doi: 10.1016/j.jcis.2012.01.053
|
| [53] |
Bode JC, Kuntsche J, Funari SS, et al. (2013) Interaction of dispersed cubic phases with blood components. Int J Pharm 448: 87-95. doi: 10.1016/j.ijpharm.2013.03.016
|
| [54] |
Azmi IDM, Wu L, Wibroe PP, et al. (2015) Modulatory effect of human plasma on the internal nanostructure and size characteristics of liquid crystalline nanocarriers. Langmuir 31: 5042-5049. doi: 10.1021/acs.langmuir.5b00830
|
| [55] |
Nilsson C, Barrios-Lopez B, Kallinen A, et al. (2013) SPECT/CT imaging of radiolabeled cubosomes and hexosomes for potential theranostic applications. Biomaterials 34: 8491-8503. doi: 10.1016/j.biomaterials.2013.07.055
|
| [56] |
Angelov B, Angelova A, Filippov SK, et al. (2014) Multicompartment lipid cubic nanoparticles with high protein upload: millisecond dynamics of formation. ACS Nano 8: 5216-5226. doi: 10.1021/nn5012946
|
| [57] |
Yaghmur A, Laggner P, Almgren M, et al. (2008) Self-assembly in monoelaidin aqueous dispersions: direct vesicles to cubosomes transition. PLoS ONE 3: e3747. doi: 10.1371/journal.pone.0003747
|
| [58] |
Rizwan SB, Dong Y-D, Boyd BJ, et al. (2007) Characterisation of bicontinuous cubic liquid crystalline systems of phytantriol and water using cryo field emission scanning electron microscopy (cryo FESEM). Micron 38:478-485. doi: 10.1016/j.micron.2006.08.003
|
| [59] |
Sagalowicz L, Acquistapace S, Watzke HJ, et al. (2007) Study of liquid crystal space groups using controlled tilting with cryogenic transmission electron microscopy. Langmuir 23: 12003-12009. doi: 10.1021/la701410n
|
| [60] |
Percec V, Wilson DA, Leowanawat P, et al. (2010) Self-assembly of Janus dendrimers into uniform dendrimersomes and other complex architectures. Science 328: 1009-1014. doi: 10.1126/science.1185547
|
| [61] |
Siegel DP, Epand RM (1997) The mechanism of lamellar-to-inverted hexagonal phase transitions in phosphatidylethanolamine: implications for membrane fusion mechanisms. Biophys J 73: 3089-3111. doi: 10.1016/S0006-3495(97)78336-X
|
| [62] |
Siegel DP, Green WJ, Talmon Y (1994) The mechanism of lamellar-to-inverted hexagonal phase transitions: a study using temperature-jump cryo-electron microscopy. Biophys J 66: 402-414. doi: 10.1016/S0006-3495(94)80790-8
|
| [63] | Ansari MJ, Kohli K, Dixit N (2008) Microemulsions as potential drug delivery systems: a review. PDA J Pharm Sci Technol 62: 66-79. |
| [64] |
Lawrence MJ, Rees GD (2000) Microemulsion-based media as novel drug delivery systems. Adv Drug Delivery Rev 45: 89-121. doi: 10.1016/S0169-409X(00)00103-4
|
| [65] | Chatzidaki MD, Mitsou E, Yaghmur A, et al. (2015) Formulation and characterization of food-grade microemulsions as carriers of natural phenolic antioxidants. Colloid Surf A Doi: 10.1016/j.colsurfa.2015.03.060. |
| [66] |
Yaghmur A., Aserin A, Garti N (2002) Phase behavior of microemulsions based on food-grade nonionic surfactants: effect of polyols and short-chain alcohols. Colloids Surf A 209:71-81. doi: 10.1016/S0927-7757(02)00168-1
|
| [67] |
Chevalier Y, Zemb T (1990) The Structure of Micelles and Microemulsions. Rep Prog Phys 53: 279-371. doi: 10.1088/0034-4885/53/3/002
|
| [68] |
Strey R (1994) Microemulsion microstructure and interfacial curvature. Colloid Polym Sci 272: 1005-1019. doi: 10.1007/BF00658900
|
| [69] |
Gradzielski M (2008) Recent developments in the characterisation of microemulsions. Curr Opin Colloid Interface Sci 13: 263-269. doi: 10.1016/j.cocis.2007.10.006
|
| [70] |
Belkoura L, Stubenrauch C, Strey R (2004) Freeze fracture direct imaging: A new freeze fracture method for specimen preparation in cryo-transmission electron microscopy. Langmuir 20: 4391-4399. doi: 10.1021/la0303411
|
| [71] |
Won YY, Brannan AK, Davis HT, et al. (2002) Cryogenic transmission electron microscopy (cryo-TEM) of micelles and vesicles formed in water by polyethylene oxide-based block copolymers. J Phys Chem B 106: 3354-3364. doi: 10.1021/jp013639d
|
| [72] |
Subramaniam S, Milne JL (2004) Three-dimensional electron microscopy at molecular resolution. Annu Rev Biophys Biomol Struct 33: 141-155. doi: 10.1146/annurev.biophys.33.110502.140339
|
| [73] |
Milne JLS, Borgnia MJ, Bartesaghi A, et al. (2013) Cryo-electron microscopy--a primer for the non-microscopist. FEBS J 280: 28-45. doi: 10.1111/febs.12078
|
| [74] |
Lengyel J, Milne JLS, Subramaniam S (2008) Electron tomography in nanoparticle imaging and analysis. Nanomedicine 3: 125-131. doi: 10.2217/17435889.3.1.125
|
| [75] |
Hoenger A, Bouchet-Marquis C (2011) Cellular tomography. Adv Protein Chem Struct Biol 82: 67-90. doi: 10.1016/B978-0-12-386507-6.00003-8
|
| [76] |
Hurbain I, Sachse M (2011) The future is cold: cryo-preparation methods for transmission electron microscopy of cells. Biol Cell 103: 405-420. doi: 10.1042/BC20110015
|
| [77] |
Milne JL, Subramaniam S (2009) Cryo-electron tomography of bacteria: progress, challenges and future prospects. Nat Rev Microbiol 7: 666-675. doi: 10.1038/nrmicro2183
|
| [78] | Tocheva EI, Li Z, Jensen GJ (2010) Electron cryotomography. Cold Spring Harb Perspect Biol 2: a003442. |
| [79] | Pierson J, Vos M, McIntosh JR, et al. (2011) Perspectives on electron cryo-tomography of vitreous cryo-sections. J Electron Microsc (Tokyo) 60 Suppl 1: S93-100. |
| [80] |
Subramaniam S, Bartesaghi A, Liu J, et al. (2007) Electron tomography of viruses. Curr Opin Struct Biol 17: 596-602. doi: 10.1016/j.sbi.2007.09.009
|
| [81] |
Yahav T, Maimon T, Grossman E, et al. (2011) Cryo-electron tomography: gaining insight into cellular processes by structural approaches. Curr Opin Struct Biol 21: 670-677. doi: 10.1016/j.sbi.2011.07.004
|
| [82] |
Wisedchaisri G, Reichow SL, Gonen T (2011) Advances in structural and functional analysis of membrane proteins by electron crystallography. Structure 19: 1381-1393. doi: 10.1016/j.str.2011.09.001
|
| [83] |
Orlova EV, Saibil HR (2011) Structural analysis of macromolecular assemblies by electron microscopy. Chem Rev 111: 7710-7748. doi: 10.1021/cr100353t
|
| [84] |
Frank J (2009) Single-particle reconstruction of biological macromolecules in electron microscopy—30 years. Q Rev Biophys 42: 139-158. doi: 10.1017/S0033583509990059
|


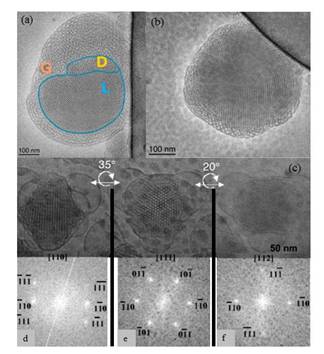


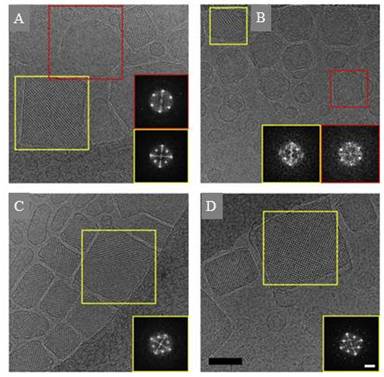
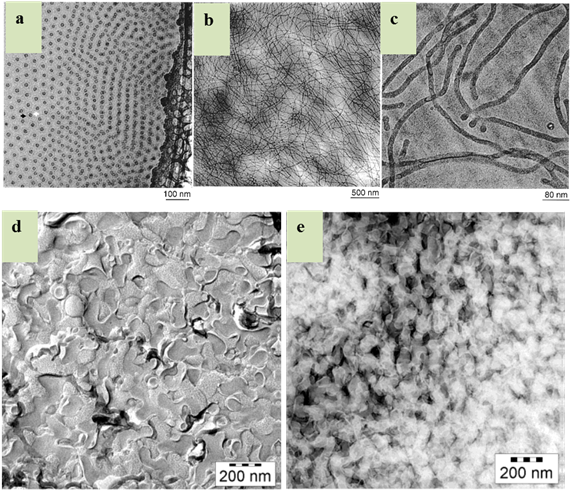
Figures(7)

Shen Helvig, Intan D. M. Azmi, Seyed M. Moghimi, Anan Yaghmur. Recent Advances in Cryo-TEM Imaging of Soft Lipid Nanoparticles[J]. AIMS Biophysics, 2015, 2(2): 116-130. doi: 10.3934/biophy.2015.2.116







 DownLoad:
DownLoad: