Citation: Caliot Cyril, Flamant Gilles. Pressurized Carbon Dioxide as Heat Transfer Fluid: In uence of Radiation on Turbulent Flow Characteristics in Pipe[J]. AIMS Energy, 2014, 1(2): 172-182. doi: 10.3934/energy.2014.2.172

| [1] | Y. Chen, W. Pridasawas, P. Lundqvist. Dynamic simulation of a solar driven carbon dioxide transcritical power system for small scale combined heat and power production, Solar Energy, 84 (2010), 1103-1110. |
| [2] | H. Chen, Y. Goswami, Stefanakos E., A review of thermodynamic cycles and working uid for the conversion of low grade heat, Renewable and Sustainable Energy Reviews, 14 (2010), 3059-3067. |
| [3] | H. Yamaguchi, X.R. Zhang, K. Fujima, M. Enomoto, N. Sawada. A solar energy powered Rankine cycle using supercritical carbon dioxide, Applied Thermal Engineering, 26 (2006), 2345-2354. |
| [4] | X.R. Zhang, H. Yamagushi, D. Uneno. Experimental study of the performance of solar Rankine system using supercritical CO2, Renewable Energy, 32 (2007), 2617-2628. |
| [5] | V. Dostal. " A supercritical carbon dioxide cycle for next generation nuclear reactors, " PhD thesis, Department of Nuclear Engineering, MIT, 2004. Available from: http://dspace.mit.edu/handle/1721.1/17746. |
| [6] | S.A. Wright, R.F. Radel, M.E. Vernon, G.E. Rochau, P.S. Pickard. Operation and analysis of a supercritical CO2 brayton cycle, Technical Report, Sandia Report SAND2010-0171, Sandia National Laboratories, 2010. Available from: http://prod.sandia.gov/techlib/access-control.cgi/2010/100171.pdf. |
| [7] | J.-E. Cha, T.-H. Lee, J.-H. Eoh, S.-H. Seong, S.-O Kim, D.-E. Kim, M. Kim, T.-W. Kim, K.-Y. Suh. Development of a supercritical CO2 brayton energy conversion system coupled with a sodium cooled fast reactor, Nuclear Engineering and Technology, 41 (2009), 1025-1044. |
| [8] | R.Z. Litwin, A.J. Zillmer, N.J. Hoffman, A.V. von Arx, D. Wait. Supercritical CO2 turbine for use in solar power plants, United States Patent US 7685820B2, 2010. |
| [9] | J. Liu, H. Chen, Y. Xu, L. Wang, C. Tan. A solar energy storage and power generation system based on supercritical carbon dioxide, Renewable Energy, 64 (2014), 43-51. |
| [10] | G. Turchi G., Z. Ma, T.W. Neises, M.J. Wagner. Thermodynamic study of advanced supercritical carbon dioxide cycles for concentrated solar power, ASME Journal of Solar Energy Engineering, 135 (2013), 041007-041013. |
| [11] | X.D. Niu, H. Yamaguchi, X.R. Zhang, Y. Iwamoto, N. Hashitani. Experimental study of heat transfer character-istics of supercritical CO2 uid in collectors of solar Rankine cycle system, Applied Thermal Engineering, 31 (2011), 1279-1285. |
| [12] | H.Y. Kim, H. Kim, J.H. Song, B.H. Cho, Y.Y. Bae. Heat transfer test in a vertical tube using CO2 at supercritical pressures, Journal of Nuclear Science and Technology, 44 (2007), 285-293. |
| [13] | R. Span, W. Wagner. A new equation of state for carbon dioxide covering the uid region from the triple-point temperature to 1100 K at pressures up to 800 MPa, Journal of Physical and Chemical Reference Data, 25 (1996), 1509-1596. |
| [14] | M.F. Modest. Radiative heat transfer. McGraw-Hill, New York, 1993. |
| [15] | G.D. Raithby, E.H. Chui. A finite-volume method for predicting a radiant heat transfer in enclosures with participating media, ASME Journal of Heat Transfer, 112 (1990), 415-423. |
| [16] | J. Taine, A. Soufiani. Gas IR radiative properties: From spectroscopic data to approximate models, Advances in Heat Transfer, 33 (1999), 295-414. |
| [17] | L.S. Rothman, I.E. Gordon, R.J. Barber, H. Dothe, R.R. Gamache, A. Goldman, V.I. Perevalov, S.A. Tashkun, J. Tennyson. HITEMP, the high-temperature molecular spectroscopic database, Journal of Quantitative Spectroscopy and Radiative Transfer, 111 (2010), 2139-2150. |
| [18] | S.A. Tashkun, V.I. Perevalov. CDSD-4000: High-resolution, high-temperature carbon dioxide spectroscopic data-bank, Journal of Quantitative Spectroscopy and Radiative Transfer, 112 (2011), 1403-1410. |
| [19] | J.-M. Hartmann, C. Boulet, D. Robert. Collisional effects on molecular spectra: Laboratory experiments and models, consequences for applications, Elsevier Sciene Ltd, 2008. |
| [20] | C. Caliot, Y. Le Maoult, M. El Hafi, G. Flamant. Remote sensing of high temperature H2O-CO2-CO mixture with a correlated k-distribution fictitious gas method and the single-mixture gas assumption, Journal of Quantitative Spectroscopy and Radiative Transfer, 102 (2006), 304-315. |
| [21] | M.Y. Perrin, J.M. Hartmann. Temperature-dependent measurements and modeling of absorption by CO2-N2 mixtures in the far line-wings of the 4.3[mu]m CO2 band, Journal of Quantitative Spectroscopy and Radiative Transfer, 42 (1989), 311-317. |
| [22] | S.V. Patankar, B.D. Spalding. A calculation procedure for heat, mass and momentum transfer in three-dimensional parabolic ows, International Journal of Heat and Mass Transfer, 15 (1972), 1787-1806. |
| [23] | B.P. Leonard, S. Mokhtari. ULTRA-SHARP nonoscillatory convection schemes for high-speed steady multidimen-sional ow, Technical Memorandum 102568 (ICOMP-90-12), NASA Lewis Research Center, 1990. |
| [24] | B. Kader. Temperature and concentration profiles in fully turbulent boundary layers, International Journal of Heat and Mass Transfer, 24 (1981), 1541-1544. |
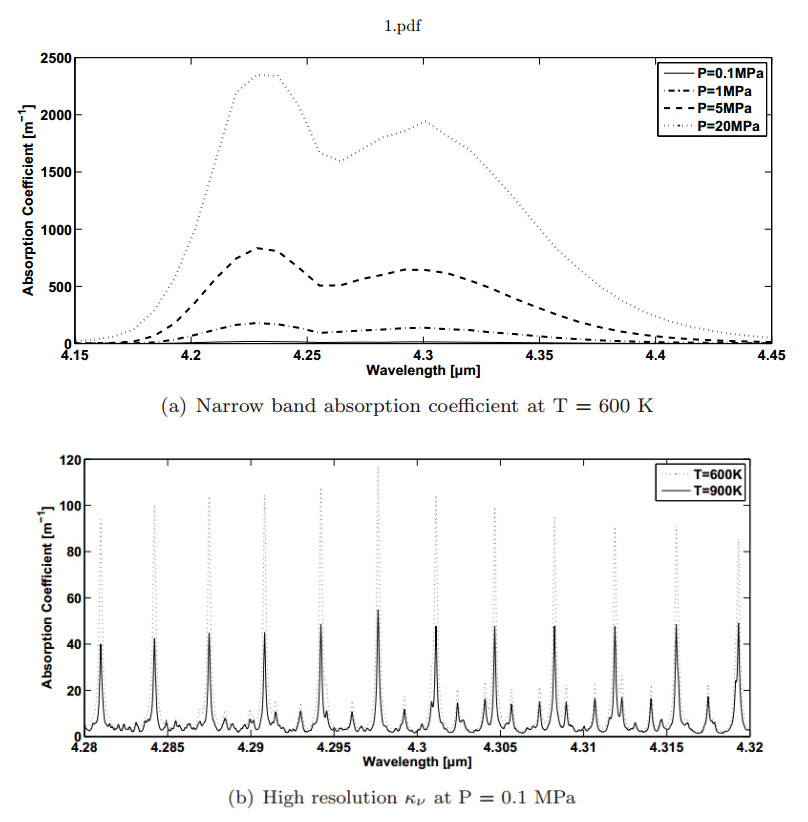
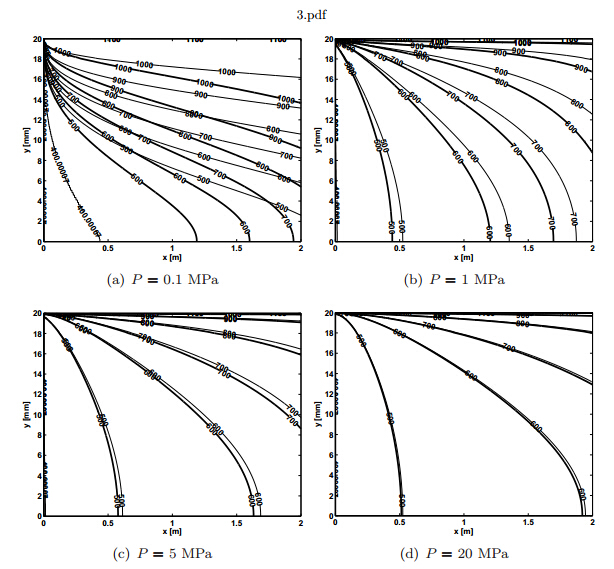
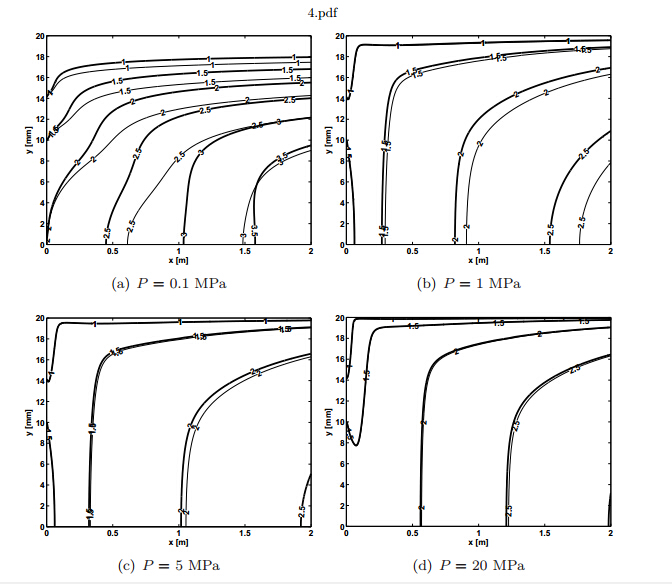
Figures(4)

Caliot Cyril, Flamant Gilles. Pressurized Carbon Dioxide as Heat Transfer Fluid: In uence of Radiation on Turbulent Flow Characteristics in Pipe[J]. AIMS Energy, 2014, 1(2): 172-182. doi: 10.3934/energy.2014.2.172







 DownLoad:
DownLoad: