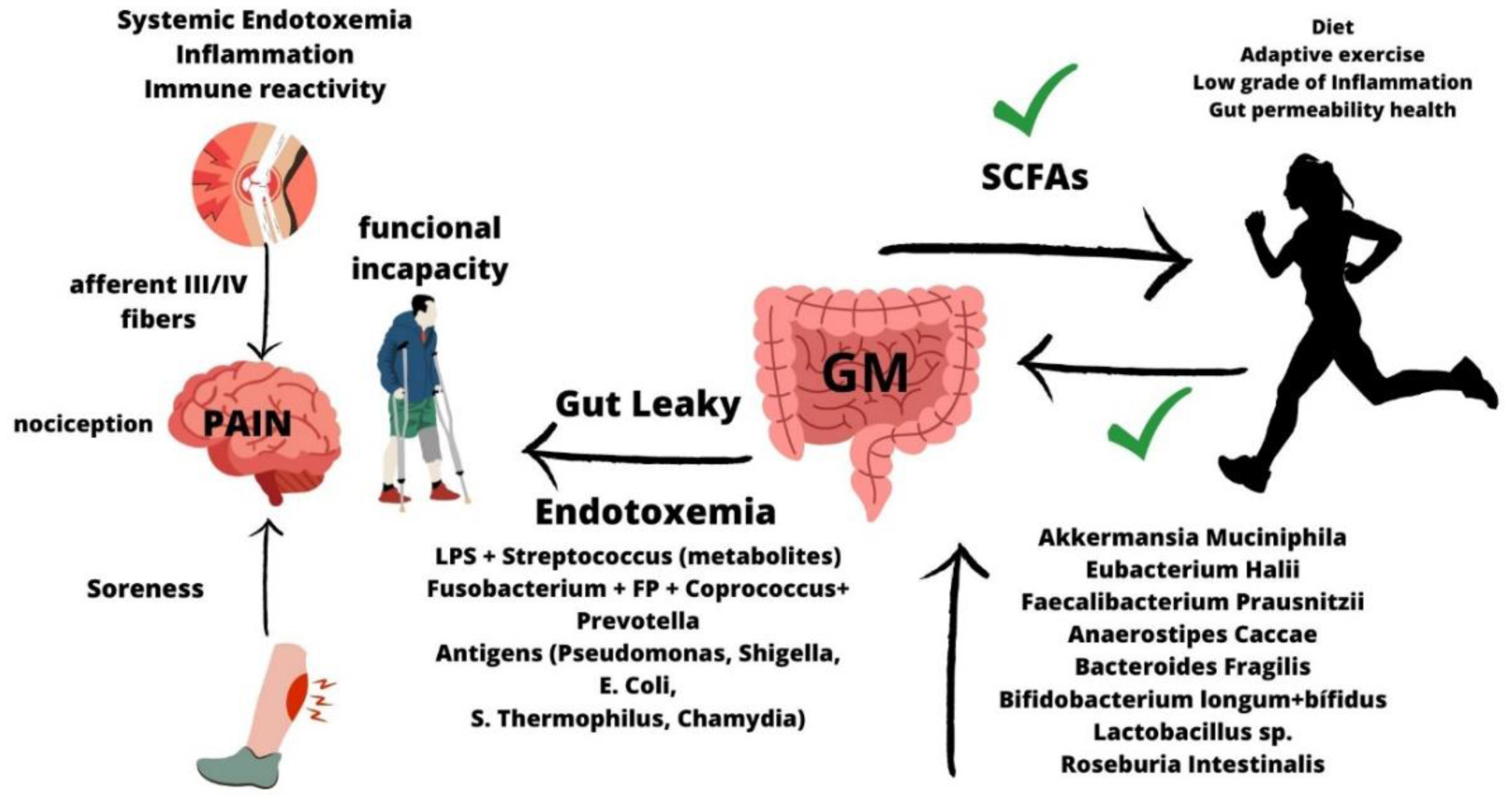
This article aims to examine the evidence on the relationship between gut microbiota (GM), leaky gut syndrome and musculoskeletal injuries. Musculoskeletal injuries can significantly impair athletic performance, overall health, and quality of life. Emerging evidence suggests that the state of the gut microbiota and the functional intestinal permeability may contribute to injury recovery. Since 2007, a growing field of research has supported the idea that GM exerts an essential role maintaining intestinal homeostasis and organic and systemic health. Leaky gut syndrome is an acquired condition where the intestinal permeability is impaired, and different bacteria and/or toxins enter in the bloodstream, thereby promoting systemic endotoxemia and chronic low-grade inflammation. This systemic condition could indirectly contribute to increased local musculoskeletal inflammation and chronificate injuries and pain, thereby reducing recovery-time and limiting sport performance. Different strategies, including a healthy diet and the intake of pre/probiotics, may contribute to improving and/or restoring gut health, thereby modulating both systemically as local inflammation and pain. Here, we sought to identify critical factors and potential strategies that could positively improve gut microbiota and intestinal health, and reduce the risk of musculoskeletal injuries and its recovery-time and pain. In conclusion, recent evidences indicate that improving gut health has indirect consequences on the musculoskeletal tissue homeostasis and recovery through the direct modulation of systemic inflammation, the immune response and the nociceptive pain.
Citation: Jesús Álvarez-Herms, Adriana González, Francisco Corbi, Iñaki Odriozola, Adrian Odriozola. Possible relationship between the gut leaky syndrome and musculoskeletal injuries: the important role of gut microbiota as indirect modulator[J]. AIMS Public Health, 2023, 10(3): 710-738. doi: 10.3934/publichealth.2023049
This article aims to examine the evidence on the relationship between gut microbiota (GM), leaky gut syndrome and musculoskeletal injuries. Musculoskeletal injuries can significantly impair athletic performance, overall health, and quality of life. Emerging evidence suggests that the state of the gut microbiota and the functional intestinal permeability may contribute to injury recovery. Since 2007, a growing field of research has supported the idea that GM exerts an essential role maintaining intestinal homeostasis and organic and systemic health. Leaky gut syndrome is an acquired condition where the intestinal permeability is impaired, and different bacteria and/or toxins enter in the bloodstream, thereby promoting systemic endotoxemia and chronic low-grade inflammation. This systemic condition could indirectly contribute to increased local musculoskeletal inflammation and chronificate injuries and pain, thereby reducing recovery-time and limiting sport performance. Different strategies, including a healthy diet and the intake of pre/probiotics, may contribute to improving and/or restoring gut health, thereby modulating both systemically as local inflammation and pain. Here, we sought to identify critical factors and potential strategies that could positively improve gut microbiota and intestinal health, and reduce the risk of musculoskeletal injuries and its recovery-time and pain. In conclusion, recent evidences indicate that improving gut health has indirect consequences on the musculoskeletal tissue homeostasis and recovery through the direct modulation of systemic inflammation, the immune response and the nociceptive pain.

| [1] |
Saxon L, Finch C, Bass S (1999) Sports participation, sports injuries and osteoarthritis: implications for prevention. Sports Med 28: 123-135. https://doi.org/10.2165/00007256-199928020-00005 
|
| [2] |
Robinson WH, Lepus CM, Wang Q, et al. (2016) Low-grade inflammation as a key mediator of the pathogenesis of osteoarthritis. Nat Rev Rheumatol 12: 580-592. https://doi.org/10.1038/nrrheum.2016.136
|
| [3] |
Chisari E, Rehak L, Khan WS, et al. (2021) Tendon healing is adversely affected by low-grade inflammation. J Orthop Surg Res 16: 700. https://doi.org/10.1186/s13018-021-02811-w
|
| [4] |
Chazaud B (2020) Inflammation and Skeletal Muscle Regeneration: Leave It to the Macrophages!. Trends Immunol 41: 481-492. https://doi.org/10.1016/j.it.2020.04.006
|
| [5] |
Silbernagel KG, Hanlon S, Sprague A (2020) Current Clinical Concepts: Conservative Management of Achilles Tendinopathy. J Athl Train 55: 438-447. https://doi.org/10.4085/1062-6050-356-19
|
| [6] |
Khan A, Khan S, Kim YS (2019) Insight into Pain Modulation: Nociceptors Sensitization and Therapeutic Targets. Curr Drug Targets 20: 775-788. https://doi.org/10.2174/1389450120666190131114244
|
| [7] |
Basbaum AI, Bautista DM, Scherrer G, et al. (2009) Cellular and molecular mechanisms of pain. Cell 139: 267-284. https://doi.org/10.1016/j.cell.2009.09.0288
|
| [8] | O'Connor PJ, Cook DB (1999) Exercise and pain: the neurobiology, measurement, and laboratory study of pain in relation to exercise in humans. Exerc Sport Sci Rev 27: 119-166. |
| [9] |
Maynard CL, Elson CO, Hatton RD, et al. (2012) Reciprocal interactions of the intestinal microbiota and immune system. Nature 489: 231-241. https://doi.org/10.1038/nature11551
|
| [10] |
Kinashi Y, Hase K (2021) Partners in Leaky Gut Syndrome: Intestinal Dysbiosis and Autoimmunity. Front Immunol 12: 673708. https://doi.org/10.3389/fimmu.2021.673708
|
| [11] |
Camilleri M (2019) Leaky gut: mechanisms, measurement and clinical implications in humans. Gut 68: 1516-1526. http://doi.org/10.1136/gutjnl-2019-318427
|
| [12] |
Ott SJ, Musfeldt M, Wenderoth DF, et al. (2004) Reduction in diversity of the colonic mucosa associated bacterial microflora in patients with active inflammatory bowel disease. Gut 53: 685-693. http://doi.org/10.1136/gut.2003.025403
|
| [13] |
Ley RE, Turnbaugh PJ, Klein S, et al. (2006) Microbial ecology: human gut microbes associated with obesity. Nature 444: 1022-1023. https://doi.org/10.1038/4441022a
|
| [14] |
Freidin MB, Stalteri MA, Wells PM, et al. (2021) An association between chronic widespread pain and the gut microbiome. Rheumatology 60: 3727-3737. https://doi.org/10.1093/rheumatology/keaa847
|
| [15] |
Puértolas-Balint F, Schroeder BO (2020) Does an Apple a Day Also Keep the Microbes Away? The Interplay Between Diet, Microbiota, and Host Defense Peptides at the Intestinal Mucosal Barrier. Frontiers in Immunology 11. https://doi.org/10.3389/fimmu.2020.01164
|
| [16] | Zhang SL, Bai L, Goel N, et al. (2017) Human and rat gut microbiome composition is maintained following sleep restriction. Proc Natl Acad Sci USA 114: E1564-E1571. https://doi.org/10.1073/pnas.1620673114 |
| [17] |
Boelsterli UA, Redinbo MR, Saitta KS (2013) Multiple NSAID-induced hits injure the small intestine: underlying mechanisms and novel strategies. Toxicol Sci 131: 654-667. https://doi.org/10.1093/toxsci/kfs310
|
| [18] |
Tristan Asensi M, Napoletano A, Sofi F, et al. (2023) Low-Grade Inflammation and Ultra-Processed Foods Consumption: A Review. Nutrients 15: 1546. https://doi.org/10.3390/nu15061546
|
| [19] |
Bischoff SC, Barbara G, Buurman W, et al. (2014) Intestinal permeability--a new target for disease prevention and therapy. BMC Gastroenterol 14: 189. https://doi.org/10.1186/s12876-014-0189-7
|
| [20] |
Sadowska-Krępa E, Rozpara M, Rzetecki A, et al. (2021) Strenuous 12-h run elevates circulating biomarkers of oxidative stress, inflammation and intestinal permeability in middle-aged amateur runners: A preliminary study. PLoS One 16: e0249183. https://doi.org/10.1371/journal.pone.0249183
|
| [21] |
Chantler S, Griffiths A, Matu J, et al. (2021) The Effects of Exercise on Indirect Markers of Gut Damage and Permeability: A Systematic Review and Meta-analysis. Sports Med 51: 113-124. https://doi.org/10.1007/s40279-020-01348-y
|
| [22] |
Mach N, Fuster-Botella D (2017) Endurance exercise and gut microbiota: A review. J Sport Health Sci 6: 179-197. https://doi.org/10.1016/j.jshs.2016.05.001
|
| [23] |
Bjørklund G, Aaseth J, Doşa MD, et al. (2019) Does diet play a role in reducing nociception related to inflammation and chronic pain?. Nutrition 66: 153-165. https://doi.org/10.1016/j.nut.2019.04.007
|
| [24] |
Chiu IM, Heesters BA, Ghasemlou N, et al. (2013) Bacteria activate sensory neurons that modulate pain and inflammation. Nature 501: 52-57. https://doi.org/10.1038/nature12479
|
| [25] |
Sibille KT, King C, Garrett TJ, et al. (2018) Omega-6: Omega-3 PUFA Ratio, Pain, Functioning, and Distress in Adults With Knee Pain. Clin J Pain 34: 182-189. https://doi.org/10.1097/AJP.0000000000000517
|
| [26] |
Jhun J, Cho K-H, Lee D-H, et al. (2021) Oral Administration of Lactobacillus rhamnosus Ameliorates the Progression of Osteoarthritis by Inhibiting Joint Pain and Inflammation. Cells 10: 1057. https://doi.org/10.3390/cells10051057
|
| [27] |
Guo R, Chen L-H, Xing C, et al. (2019) Pain regulation by gut microbiota: molecular mechanisms and therapeutic potential. Br J Anaesth 123: 637-654. https://doi.org/10.1016/j.bja.2019.07.026
|
| [28] |
Shing CM, Peake JM, Lim CL, et al. (2014) Effects of probiotics supplementation on gastrointestinal permeability, inflammation and exercise performance in the heat. Eur J Appl Physiol 114: 93-103. https://doi.org/10.1007/s00421-013-2748-y
|
| [29] |
Yeh YJ, Law LYL, Lim CL (2013) Gastrointestinal response and endotoxemia during intense exercise in hot and cool environments. Eur J Appl Physiol 113: 1575-1583. https://doi.org/10.1007/s00421-013-2587-x
|
| [30] |
de Oliveira EP, Burini RC, Jeukendrup A (2014) Gastrointestinal complaints during exercise: prevalence, etiology, and nutritional recommendations. Sports Med 44: 79-85. https://doi.org/10.1007/s40279-014-0153-2
|
| [31] |
Karhu E, Forsgård RA, Alanko L, et al. (2017) Exercise and gastrointestinal symptoms: running-induced changes in intestinal permeability and markers of gastrointestinal function in asymptomatic and symptomatic runners. Eur J Appl Physiol 117: 2519-2526. https://doi.org/10.1007/s00421-017-3739-1
|
| [32] |
March DS, Marchbank T, Playford RJ, et al. (2017) Intestinal fatty acid-binding protein and gut permeability responses to exercise. Eur J Appl Physiol 117: 931-941. https://doi.org/10.1007/s00421-017-3582-4
|
| [33] |
Snipe RMJ, Khoo A, Kitic CM, et al. (2018) The impact of exertional-heat stress on gastrointestinal integrity, gastrointestinal symptoms, systemic endotoxin and cytokine profile. Eur J Appl Physiol 118: 389-400. https://doi.org/10.1007/s00421-017-3781-z
|
| [34] |
Zuhl MN, Lanphere KR, Kravitz L, et al. (2014) Effects of oral glutamine supplementation on exercise-induced gastrointestinal permeability and tight junction protein expression. J Appl Physiol 116: 183-191. https://doi.org/10.1152/japplphysiol.00646.2013
|
| [35] |
Lamprecht M, Bogner S, Schippinger G, et al. (2012) Probiotic supplementation affects markers of intestinal barrier, oxidation, and inflammation in trained men; a randomized, double-blinded, placebo-controlled trial. J Int Soc Sports Nutr 9: 45. https://doi.org/10.1186/1550-2783-9-45
|
| [36] |
van Wijck K, Lenaerts K, Grootjans J, et al. (2012) Physiology and pathophysiology of splanchnic hypoperfusion and intestinal injury during exercise: strategies for evaluation and prevention. Am J Physiol Gastrointest Liver Physiol 303: G155-G168. https://doi.org/10.1152/ajpgi.00066.2012
|
| [37] |
Shah YM (2016) The role of hypoxia in intestinal inflammation. Molecular and Cellular Pediatrics 3: 1-5. https://doi.org/10.1186/s40348-016-0030-1
|
| [38] |
Pires W, Veneroso CE, Wanner SP, et al. (2017) Association Between Exercise-Induced Hyperthermia and Intestinal Permeability: A Systematic Review. Sports Med 47: 1389-1403. https://doi.org/10.1007/s40279-016-0654-2
|
| [39] |
Lambert GP, Lang J, Bull A, et al. (2008) Fluid restriction during running increases GI permeability. Int J Sports Med 29: 194-198. https://doi.org/0.1055/s-2007-965163
|
| [40] |
Vitetta L, Coulson S, Linnane AW, et al. (2013) The gastrointestinal microbiome and musculoskeletal diseases: a beneficial role for probiotics and prebiotics. Pathogens 2: 606-626. https://doi.org/10.3390/pathogens2040606
|
| [41] |
Li C, Li Y, Wang N, et al. (2022) Intestinal Permeability Associated with the Loss of Skeletal Muscle Strength in Middle-Aged and Older Adults in Rural Area of Beijing, China. Healthcare (Basel) 10: 1100. https://doi.org/10.3390/healthcare10061100
|
| [42] |
Ohlsson C, Sjögren K (2015) Effects of the gut microbiota on bone mass. Trends Endocrinol Metab 26: 69-74. https://doi.org/10.1016/j.tem.2014.11.004
|
| [43] |
Boer CG, Radjabzadeh D, Medina-Gomez C, et al. (2019) Intestinal microbiome composition and its relation to joint pain and inflammation. Nat Commun 10: 4881. https://doi.org/10.1038/s41467-019-12873-4
|
| [44] |
Schwellnus M, Soligard T, Alonso J-M, et al. (2016) How much is too much? (Part 2) International Olympic Committee consensus statement on load in sport and risk of illness. Br J Sports Med 50: 1043-1052. http://doi.org/10.1136/bjsports-2016-096572
|
| [45] |
Korpela K, de Vos WM (2018) Early life colonization of the human gut: microbes matter everywhere. Curr Opin Microbiol 44: 70-78. https://doi.org/10.1016/j.mib.2018.06.003
|
| [46] |
Vijay A, Valdes AM (2022) Role of the gut microbiome in chronic diseases: a narrative review. Eur J Clin Nutr 76: 489-501. https://doi.org/10.1038/s41430-021-00991-6
|
| [47] |
Das B, Nair GB (2019) Homeostasis and dysbiosis of the gut microbiome in health and disease. J Biosci 44: 117. https://doi.org/10.1007/s12038-019-9926-y
|
| [48] |
Boer CG, Radjabzadeh D, Medina-Gomez C, et al. (2019) Intestinal microbiome composition and its relation to joint pain and inflammation. Nat Commun 10: 4881. https://doi.org/10.1038/s41467-019-12873-4
|
| [49] |
Brenner D, Shorten GD, O'Mahony SM (2021) Postoperative pain and the gut microbiome. Neurobiol Pain 10: 100070. https://doi.org/10.1016/j.ynpai.2021.100070
|
| [50] |
Morrison SA, Cheung SS, Cotter JD (2014) Bovine colostrum, training status, and gastrointestinal permeability during exercise in the heat: a placebo-controlled double-blind study. Appl Physiol Nutr Metab 39: 1070-1082. https://doi.org/10.1139/apnm-2013-0583
|
| [51] |
Miquel S, Martín R, Lashermes A, et al. (2016) Anti-nociceptive effect of Faecalibacterium prausnitzii in non-inflammatory IBS-like models. Sci Rep 6: 19399. https://doi.org/10.1038/srep19399
|
| [52] |
Tonelli Enrico V, Vo N, Methe B, et al. (2022) An unexpected connection: A narrative review of the associations between Gut Microbiome and Musculoskeletal Pain. Eur Spine J 31: 3603-3615. https://doi.org/10.1007/s00586-022-07429-y
|
| [53] |
Hiippala K, Jouhten H, Ronkainen A, et al. (2018) The Potential of Gut Commensals in Reinforcing Intestinal Barrier Function and Alleviating Inflammation. Nutrients 10: 988. https://doi.org/10.3390/nu10080988
|
| [54] |
Neish AS (2009) Microbes in gastrointestinal health and disease. Gastroenterology 136: 65-80. https://doi.org/10.1053/j.gastro.2008.10.080
|
| [55] |
Turner JR (2009) Intestinal mucosal barrier function in health and disease. Nat Rev Immunol 9: 799-809. https://doi.org/10.1038/nri2653
|
| [56] |
Cone RA (2009) Barrier properties of mucus. Adv Drug Deliv Rev 61: 75-85. https://doi.org/10.1016/j.addr.2008.09.008
|
| [57] |
Suzuki T (2013) Regulation of intestinal epithelial permeability by tight junctions. Cell Mol Life Sci 70: 631-659. https://doi.org/10.1007/s00018-012-1070-x
|
| [58] |
O'Hara AM, Shanahan F (2006) The gut flora as a forgotten organ. EMBO Rep 7: 688-693. https://doi.org/10.1038/sj.embor.7400731
|
| [59] |
Belzer C, Chia LW, Aalvink S, et al. (2017) Microbial Metabolic Networks at the Mucus Layer Lead to Diet-Independent Butyrate and Vitamin B12 Production by Intestinal Symbionts. MBio 8: e00770-17. https://doi.org/10.1128/mbio.00770-17
|
| [60] |
Strandwitz P (2018) Neurotransmitter modulation by the gut microbiota. Brain Research 1693: 128-133. https://doi.org/10.1016/j.brainres.2018.03.015
|
| [61] |
Suez J, Korem T, Zeevi D, et al. (2014) Artificial sweeteners induce glucose intolerance by altering the gut microbiota. Nature 514: 181-186. https://doi.org/10.1038/nature13793
|
| [62] |
Martarelli D, Verdenelli MC, Scuri S, et al. (2011) Effect of a probiotic intake on oxidant and antioxidant parameters in plasma of athletes during intense exercise training. Curr Microbiol 62: 1689-1696. https://doi.org/10.1007/s00284-011-9915-3
|
| [63] |
Spyropoulos BG, Misiakos EP, Fotiadis C, et al. (2011) Antioxidant properties of probiotics and their protective effects in the pathogenesis of radiation-induced enteritis and colitis. Dig Dis Sci 56: 285-294. https://doi.org/10.3390/nu9050521
|
| [64] |
Rinninella E, Cintoni M, Raoul P, et al. (2019) Food Components and Dietary Habits: Keys for a Healthy Gut Microbiota Composition. Nutrients 11: 2393. https://doi.org/10.3390/nu11102393
|
| [65] |
Lin B, Wang Y, Zhang P, et al. (2020) Gut microbiota regulates neuropathic pain: potential mechanisms and therapeutic strategy. J Headache Pain 21: 103. https://doi.org/10.1186/s10194-020-01170-x
|
| [66] |
Huang Z, Kraus VB (2016) Does lipopolysaccharide-mediated inflammation have a role in OA?. Nat Rev Rheumatol 12: 123-129. https://doi.org/10.1038/nrrheum.2015.158
|
| [67] |
Schumann RR, Leong SR, Flaggs GW, et al. (1990) Structure and function of lipopolysaccharide binding protein. Science 249: 1429-1431. https://doi.org/10.1126/science.2402637
|
| [68] | Adamu B, Sani MU, Abdu A (2006) Physical exercise and health: a review. Niger J Med 15: 190-196. https://doi.org/10.4314/njm.v15i3.37214 |
| [69] |
Chéron C, Le Scanff C, Leboeuf-Yde C (2017) Association between sports type and overuse injuries of extremities in adults: a systematic review. Chiropr Man Therap 25: 4. https://doi.org/10.1186/s12998-017-0135-1
|
| [70] |
Furman D, Campisi J, Verdin E, et al. (2019) Chronic inflammation in the etiology of disease across the life span. Nat Med 25: 1822-1832. https://doi.org/10.1038/s41591-019-0675-0
|
| [71] |
Medzhitov R (2008) Origin and physiological roles of inflammation. Nature 454: 428-435. https://doi.org/10.1038/nature07201
|
| [72] |
Cornish SM, Chilibeck PD, Candow DG (2020) Potential Importance of Immune System Response to Exercise on Aging Muscle and Bone. Curr Osteoporos Rep 18: 350-356. https://doi.org/10.1007/s11914-020-00596-1
|
| [73] |
Picca A, Fanelli F, Calvani R, et al. (2018) Gut Dysbiosis and Muscle Aging: Searching for Novel Targets against Sarcopenia. Mediators Inflamm 2018: 7026198. https://doi.org/10.1155/2018/7026198
|
| [74] |
Pérez-Baos S, Prieto-Potin I, Román-Blas JA, et al. (2018) Mediators and Patterns of Muscle Loss in Chronic Systemic Inflammation. Frontiers in Physiology 9. https://doi.org/10.3389/fphys.2018.00409
|
| [75] |
Zhou WBS, Meng J, Zhang J (2021) Does Low Grade Systemic Inflammation Have a Role in Chronic Pain?. Front Mol Neurosci 14: 785214. https://doi.org/10.3389/fnmol.2021.785214
|
| [76] |
Gao HGL, Fisher PW, Lambi AG, et al. (2013) Increased serum and musculotendinous fibrogenic proteins following persistent low-grade inflammation in a rat model of long-term upper extremity overuse. PLoS One 8: e71875. https://doi.org/10.1371/journal.pone.0071875
|
| [77] | Dessem D, Lovering RM (2011) Repeated muscle injury as a presumptive trigger for chronic masticatory muscle pain. Pain Res Treat 2011: 647967. https://doi.org/10.1155/2011/647967 |
| [78] |
Conboy IM, Conboy MJ, Wagers AJ, et al. (2005) Rejuvenation of aged progenitor cells by exposure to a young systemic environment. Nature 433: 760-764. https://doi.org/10.1038/nature03260
|
| [79] |
Franceschi C, Campisi J (2014) Chronic inflammation (inflammaging) and its potential contribution to age-associated diseases. J Gerontol A Biol Sci Med Sci 69: S4-S9. https://doi.org/10.1093/gerona/glu057
|
| [80] |
Remels AH, Gosker HR, van der Velden J, et al. (2007) Systemic inflammation and skeletal muscle dysfunction in chronic obstructive pulmonary disease: state of the art and novel insights in regulation of muscle plasticity. Clin Chest Med 28: 537-552. https://doi.org/10.1016/j.ccm.2007.06.003
|
| [81] |
Bonaldo P, Sandri M (2013) Cellular and molecular mechanisms of muscle atrophy. Dis Model Mech 6: 25-39. https://doi.org/10.1242/dmm.010389
|
| [82] |
de Sire R, Rizzatti G, Ingravalle F, et al. (2018) Skeletal muscle-gut axis: emerging mechanisms of sarcopenia for intestinal and extra intestinal diseases. Minerva Gastroenterol Dietol 64: 351-362. https://doi.org/10.23736/s1121-421x.18.02511-4
|
| [83] |
Li R, Boer CG, Oei L, et al. (2021) The Gut Microbiome: a New Frontier in Musculoskeletal Research. Curr Osteoporos Rep 19: 347-357. https://doi.org/10.1007/s11914-021-00675-x
|
| [84] |
Grosicki GJ, Fielding RA, Lustgarten MS (2018) Gut Microbiota Contribute to Age-Related Changes in Skeletal Muscle Size, Composition, and Function: Biological Basis for a Gut-Muscle Axis. Calcif Tissue Int 102: 433-442. https://doi.org/10.1007/s00223-017-0345-5
|
| [85] |
Huang W-C, Chen Y-H, Chuang H-L, et al. (2019) Investigation of the Effects of Microbiota on Exercise Physiological Adaption, Performance, and Energy Utilization Using a Gnotobiotic Animal Model. Front Microbiol 10: 1906. https://doi.org/10.3389/fmicb.2019.01906
|
| [86] |
Nay K, Jollet M, Goustard B, et al. (2019) Gut bacteria are critical for optimal muscle function: a potential link with glucose homeostasis. Am J Physiol Endocrinol Metab 317: E158-E171. https://doi.org/10.1152/ajpendo.00521.2018
|
| [87] |
Fielding RA, Reeves AR, Jasuja R, et al. (2019) Muscle strength is increased in mice that are colonized with microbiota from high-functioning older adults. Exp Gerontol 127: 110722. https://doi.org/10.1016/j.exger.2019.110722
|
| [88] |
Tidball JG (2005) Inflammatory processes in muscle injury and repair. Am J Physiol Regul Integr Comp Physiol 288: R345-R353. https://doi.org/10.1152/ajpregu.00454.2004
|
| [89] |
Webster JM, Kempen LJAP, Hardy RS, et al. (2020) Inflammation and Skeletal Muscle Wasting During Cachexia. Front Physiol 11: 597675. https://doi.org/10.3389/fphys.2020.597675
|
| [90] |
Przybyla B, Gurley C, Harvey JF, et al. (2006) Aging alters macrophage properties in human skeletal muscle both at rest and in response to acute resistance exercise. Exp Gerontol 41: 320-327. https://doi.org/10.1016/j.exger.2005.12.007
|
| [91] | Alvarez B, Quinn LS, Busquets S, et al. (2001) Direct effects of tumor necrosis factor alpha (TNF-alpha) on murine skeletal muscle cell lines. Bimodal effects on protein metabolism. Eur Cytokine Netw 12: 399-410. |
| [92] |
Sin DD, Reid WD (2008) Is inflammation good, bad or irrelevant for skeletal muscles in COPD?. Thorax 63: 95-96. https://doi.org/10.1136/thx.2007.088575
|
| [93] |
Pelosi L, Giacinti C, Nardis C, et al. (2007) Local expression of IGF-1 accelerates muscle regeneration by rapidly modulating inflammatory cytokines and chemokines. FASEB J 21: 1393-1402. https://doi.org/10.1096/fj.06-7690com
|
| [94] |
Yan J, Charles JF (2018) Gut Microbiota and IGF-1. Calcif Tissue Int 102: 406-414. https://doi.org/10.1007/s00223-018-0395-3
|
| [95] |
Giron M, Thomas M, Dardevet D, et al. (2022) Gut microbes and muscle function: can probiotics make our muscles stronger?. J Cachexia Sarcopenia Muscle 13: 1460-1476. https://doi.org/10.1002/jcsm.12964
|
| [96] |
Beavers KM, Brinkley TE, Nicklas BJ (2010) Effect of exercise training on chronic inflammation. Clin Chim Acta 411: 785-793. https://doi.org/10.1016/j.cca.2010.02.069
|
| [97] |
Cani PD, Amar J, Iglesias MA, et al. (2007) Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes 56: 1761-1772. https://doi.org/10.2337/db06-1491
|
| [98] | Karl JP, Hatch AM, Arcidiacono SM, et al. (2018) Effects of Psychological, Environmental and Physical Stressors on the Gut Microbiota. Frontiers in Microbiology 9. https://doi.org/10.3389/fmicb.2018.02013 |
| [99] |
Boets E, Gomand SV, Deroover L, et al. (2017) Systemic availability and metabolism of colonic-derived short-chain fatty acids in healthy subjects: a stable isotope study. J Physiol 595: 541-555. https://doi.org/10.1113/JP272613
|
| [100] |
Kannus P (1997) Tendons--a source of major concern in competitive and recreational athletes. Scand J Med Sci Sports 7: 53-54. https://doi.org/10.1111/j.1600-0838.1997.tb00118.x
|
| [101] | Lui PPY, Yung PSH (2021) Inflammatory mechanisms linking obesity and tendinopathy. J Orthop Transl 31: 80-90. https://doi.org/10.1016/j.jot.2021.10.003 |
| [102] |
Varela-Eirin M, Loureiro J, Fonseca E, et al. (2018) Cartilage regeneration and ageing: Targeting cellular plasticity in osteoarthritis. Ageing Res Rev 42: 56-71. https://doi.org/10.1016/j.arr.2017.12.006
|
| [103] |
Dietrich F, Hammerman M, Blomgran P, et al. (2017) Effect of platelet-rich plasma on rat Achilles tendon healing is related to microbiota. Acta Orthop 88: 463. https://doi.org/10.1080/17453674.2017.1338417
|
| [104] |
Dietrich-Zagonel F, Hammerman M, Eliasson P, et al. (2020) Response to mechanical loading in rat Achilles tendon healing is influenced by the microbiome. PLoS One 15: e0229908. https://doi.org/10.1371/journal.pone.0229908
|
| [105] |
Dietrich-Zagonel F, Hammerman M, Tätting L, et al. (2018) Stimulation of Tendon Healing With Delayed Dexamethasone Treatment Is Modified by the Microbiome. Am J Sports Med 46: 3281-3287. https://doi.org/10.1177/0363546518799442
|
| [106] |
Barbour KE (2017) Vital Signs: Prevalence of Doctor-Diagnosed Arthritis and Arthritis-Attributable Activity Limitation — United States, 2013–2015. MMWR Morb Mortal Wkly Rep 66. http://doi.org/10.15585/mmwr.mm6609e1
|
| [107] |
Korotkyi O, Kyriachenko Y, Kobyliak N, et al. (2020) Crosstalk between gut microbiota and osteoarthritis: A critical view. J Funct Foods 68: 103904. https://doi.org/10.1016/j.jff.2020.103904
|
| [108] |
Metcalfe D, Harte AL, Aletrari MO, et al. (2012) Does endotoxaemia contribute to osteoarthritis in obese patients?. Clin Sci (Lond) 123: 627-634. https://doi.org/10.1042/CS20120073
|
| [109] |
de Sire A, de Sire R, Petito V, et al. (2020) Gut-Joint Axis: The Role of Physical Exercise on Gut Microbiota Modulation in Older People with Osteoarthritis. Nutrients 12: 574. https://doi.org/10.3390/nu12020574
|
| [110] |
Favazzo LJ, Hendesi H, Villani DA, et al. (2020) The gut microbiome-joint connection: implications in osteoarthritis. Curr Opin Rheumatol 32: 92-101. https://doi.org/10.1097/BOR.0000000000000681
|
| [111] |
Huang Z, Chen J, Li B, et al. (2020) Faecal microbiota transplantation from metabolically compromised human donors accelerates osteoarthritis in mice. Ann Rheum Dis 79: 646-656. http://doi.org/10.1136/annrheumdis-2019-216471
|
| [112] |
Andoh A, Nishida A, Takahashi K, et al. (2016) Comparison of the gut microbial community between obese and lean peoples using 16S gene sequencing in a Japanese population. J Clin Biochem Nutr 59: 65-70. https://doi.org/10.3164/jcbn.15-152
|
| [113] |
Liu R, Hong J, Xu X, et al. (2017) Gut microbiome and serum metabolome alterations in obesity and after weight-loss intervention. Nat Med 23: 859-868. https://doi.org/10.1038/nm.4358
|
| [114] |
Milani C, Duranti S, Bottacini F, et al. (2017) The First Microbial Colonizers of the Human Gut: Composition, Activities, and Health Implications of the Infant Gut Microbiota. Microbiol Mol Biol Rev 81: e00036-17. https://doi.org/10.1128/MMBR.00036-17
|
| [115] |
Xiao S, Fei N, Pang X, et al. (2014) A gut microbiota-targeted dietary intervention for amelioration of chronic inflammation underlying metabolic syndrome. FEMS Microbiol Ecol 87: 357-367. https://doi.org/10.1111/1574-6941.12228
|
| [116] |
Wei J, Zhang C, Zhang Y, et al. (2021) Association Between Gut Microbiota and Symptomatic Hand Osteoarthritis: Data From the Xiangya Osteoarthritis Study. Arthritis Rheumatol 73: 1656-1662. https://doi.org/10.1002/art.41729
|
| [117] |
Coulson S, Butt H, Vecchio P, et al. (2013) Green-lipped mussel extract (Perna canaliculus) and glucosamine sulphate in patients with knee osteoarthritis: therapeutic efficacy and effects on gastrointestinal microbiota profiles. Inflammopharmacology 21: 79-90. https://doi.org/10.1007/s10787-012-0146-4
|
| [118] |
Chen J, Wang A, Wang Q (2021) Dysbiosis of the gut microbiome is a risk factor for osteoarthritis in older female adults: a case control study. BMC Bioinformatics 22: 299. https://doi.org/10.1186/s12859-021-04199-0
|
| [119] |
Kempsell KE, Cox CJ, Hurle M, et al. (2000) Reverse transcriptase-PCR analysis of bacterial rRNA for detection and characterization of bacterial species in arthritis synovial tissue. Infect Immun 68: 6012-6026. https://doi.org/10.1128/iai.68.10.6012-6026.2000
|
| [120] |
Silverstein FE, Faich G, Goldstein JL, et al. (2000) Gastrointestinal toxicity with celecoxib vs nonsteroidal anti-inflammatory drugs for osteoarthritis and rheumatoid arthritis: the CLASS study: A randomized controlled trial. Celecoxib Long-term Arthritis Safety Study. JAMA 284: 1247-1255. https://doi.org/10.1001/jama.284.10.1247
|
| [121] |
Guan Z, Jia J, Zhang C, et al. (2020) Gut microbiome dysbiosis alleviates the progression of osteoarthritis in mice. Clin Sci (Lond) 134: 3159-3174. http://doi.org/10.2139/ssrn.3696788
|
| [122] |
Schott EM, Farnsworth CW, Grier A, et al. (2018) Targeting the gut microbiome to treat the osteoarthritis of obesity. JCI Insight 3: e95997. https://doi.org/10.1172/jci.insight.95997
|
| [123] |
Rios JL, Bomhof MR, Reimer RA, et al. (2019) Protective effect of prebiotic and exercise intervention on knee health in a rat model of diet-induced obesity. Sci Rep 9: 3893. https://doi.org/10.1038/s41598-019-40601-x
|
| [124] |
So J-S, Song M-K, Kwon H-K, et al. (2011) Lactobacillus casei enhances type II collagen/glucosamine-mediated suppression of inflammatory responses in experimental osteoarthritis. Life Sci 88: 358-366. https://doi.org/10.1016/j.lfs.2010.12.013
|
| [125] |
Mandel DR, Eichas K, Holmes J (2010) Bacillus coagulans: a viable adjunct therapy for relieving symptoms of rheumatoid arthritis according to a randomized, controlled trial. BMC Complement Altern Med 10: 1. https://doi.org/10.1186/1472-6882-10-1
|
| [126] |
O'Keefe RJ, Mao J (2011) Bone tissue engineering and regeneration: from discovery to the clinic--an overview. Tissue Eng Part B Rev 17: 389-392. https://doi.org/10.1089/ten.teb.2011.0475
|
| [127] |
Loi F, Córdova LA, Pajarinen J, et al. (2016) Inflammation, fracture and bone repair. Bone 86: 119-130. https://doi.org/10.1016/j.bone.2016.02.020
|
| [128] |
Harrast MA, Colonno D (2010) Stress fractures in runners. Clin Sports Med 29: 399-416. https://doi.org/10.1016/j.csm.2010.03.001
|
| [129] |
Goolsby MA, Boniquit N (2016) Bone Health in Athletes. Sports Health 9: 108-117. https://doi.org/10.1177/1941738116677732
|
| [130] |
Zhernakova A, Kurilshikov A, Bonder MJ, et al. (2016) Population-based metagenomics analysis reveals markers for gut microbiome composition and diversity. Science 352: 565-569. https://doi.org/10.1126/science.aad3369
|
| [131] |
Horowitz MC (1993) Cytokines and estrogen in bone: anti-osteoporotic effects. Science 260: 626-627. https://doi.org/10.1126/science.8480174
|
| [132] |
Srutkova D, Schwarzer M, Hudcovic T, et al. (2015) Bifidobacterium longum CCM 7952 Promotes Epithelial Barrier Function and Prevents Acute DSS-Induced Colitis in Strictly Strain-Specific Manner. PLoS One 10: e0134050. https://doi.org/10.1371/journal.pone.0134050
|
| [133] |
Schepper JD, Collins F, Rios-Arce ND, et al. (2020) Involvement of the Gut Microbiota and Barrier Function in Glucocorticoid-Induced Osteoporosis. J Bone Miner Res 35: 801-820. https://doi.org/10.1002/jbmr.3947
|
| [134] |
Collins FL, Irwin R, Bierhalter H, et al. (2016) Lactobacillus reuteri 6475 Increases Bone Density in Intact Females Only under an Inflammatory Setting. PLoS One 11: e0153180. https://doi.org/10.1371/journal.pone.0153180
|
| [135] |
Pacifici R (2018) Bone Remodeling and the Microbiome. Cold Spring Harb Perspect Med 8: a031203. https://doi.org/10.1101/cshperspect.a031203
|
| [136] |
Willems HME, van den Heuvel EGHM, Schoemaker RJW, et al. (2017) Diet and Exercise: a Match Made in Bone. Curr Osteoporos Rep 15: 555-563. https://doi.org/10.1007/s11914-017-0406-8
|
| [137] |
Zhong X, Zhang F, Yin X, et al. (2021) Bone Homeostasis and Gut Microbial-Dependent Signaling Pathways. J Microbiol Biotechnol 31: 765-774. https://doi.org/10.4014/jmb.2104.04016
|
| [138] | Yan J, Herzog JW, Tsang K, et al. (2016) Gut microbiota induce IGF-1 and promote bone formation and growth. Proc Natl Acad Sci USA 113: E7554-E7563. https://doi.org/10.1073/pnas.1607235113 |
| [139] |
Rettedal EA, Ilesanmi-Oyelere BL, Roy NC, et al. (2021) The Gut Microbiome Is Altered in Postmenopausal Women With Osteoporosis and Osteopenia. JBMR Plus 5: e10452. https://doi.org/10.1002/jbm4.10452
|
| [140] |
Li J-Y, Chassaing B, Tyagi AM, et al. (2016) Sex steroid deficiency-associated bone loss is microbiota dependent and prevented by probiotics. J Clin Invest 126: 2049-2063. https://doi.org/10.1172/JCI86062
|
| [141] |
Ishii S, Cauley JA, Greendale GA, et al. (2013) C-reactive protein, bone strength, and nine-year fracture risk: data from the Study of Women's Health Across the Nation (SWAN). J Bone Miner Res 28: 1688-1698. https://doi.org/10.1002/jbmr.1915
|
| [142] |
Reinke S, Geissler S, Taylor WR, et al. (2013) Terminally differentiated CD8+ T cells negatively affect bone regeneration in humans. Sci Transl Med 5: 177ra36. https://doi.org/10.1126/scitranslmed.3004754
|
| [143] |
Rechardt M, Shiri R, Karppinen J, et al. (2010) Lifestyle and metabolic factors in relation to shoulder pain and rotator cuff tendinitis: a population-based study. BMC Musculoskelet Disord 11: 165. https://doi.org/10.1186/1471-2474-11-165
|
| [144] |
Kulecka M, Fraczek B, Mikula M, et al. (2020) The composition and richness of the gut microbiota differentiate the top Polish endurance athletes from sedentary controls. Gut Microbes 11: 1374-1384. https://doi.org/10.1080/19490976.2020.1758009
|
| [145] |
Liang R, Zhang S, Peng X, et al. (2019) Characteristics of the gut microbiota in professional martial arts athletes: A comparison between different competition levels. PLoS One 14: e0226240. https://doi.org/10.1371/journal.pone.0226240
|
| [146] |
Hughes RL, Holscher HD (2021) Fueling Gut Microbes: A Review of the Interaction between Diet, Exercise, and the Gut Microbiota in Athletes. Adv Nutr 12: 2190-2215. https://doi.org/10.1093/advances/nmab077
|
| [147] |
Ghosh TS, Shanahan F, O'Toole PW (2022) The gut microbiome as a modulator of healthy ageing. Nat Rev Gastroenterol Hepatol 19: 565-584. https://doi.org/10.1038/s41575-022-00605-x
|
| [148] |
Mohr AE, Jäger R, Carpenter KC, et al. (2020) The athletic gut microbiota. J Int Soc Sports Nutr 17: 24. https://doi.org/10.1186/s12970-020-00353-w
|
| [149] |
Luo M-J, Rao S-S, Tan Y-J, et al. (2020) Fasting before or after wound injury accelerates wound healing through the activation of pro-angiogenic SMOC1 and SCG2. Theranostics 10: 3779-3792.
|
| [150] |
Patterson RE, Sears DD (2017) Metabolic Effects of Intermittent Fasting. Annu Rev Nutr 37: 371-393. https://doi.org/10.1146/annurev-nutr-071816-064634
|
| [151] |
Munukka E, Rintala A, Toivonen R, et al. (2017) Faecalibacterium prausnitzii treatment improves hepatic health and reduces adipose tissue inflammation in high-fat fed mice. ISME J 11: 1667-1679. https://doi.org/10.1038/ismej.2017.24
|
| [152] |
Li B, Evivie SE, Lu J, et al. (2018) Lactobacillus helveticus KLDS1.8701 alleviates d-galactose-induced aging by regulating Nrf-2 and gut microbiota in mice. Food Funct 9: 6586-6598. https://doi.org/10.1039/C8FO01768A
|
| [153] |
Bindels LB, Beck R, Schakman O, et al. (2012) Restoring specific lactobacilli levels decreases inflammation and muscle atrophy markers in an acute leukemia mouse model. PLoS One 7: e37971. https://doi.org/10.1371/journal.pone.0037971
|
| [154] |
Finamore A, Ambra R, Nobili F, et al. (2018) Redox Role of Lactobacillus casei Shirota Against the Cellular Damage Induced by 2,2′-Azobis (2-Amidinopropane) Dihydrochloride-Induced Oxidative and Inflammatory Stress in Enterocytes-Like Epithelial Cells. Front Immunol 9: 1131. https://doi.org/10.3389/fimmu.2018.01131
|
| [155] |
Lin R, Liu W, Piao M, et al. (2017) A review of the relationship between the gut microbiota and amino acid metabolism. Amino Acids 49: 2083-2090. https://doi.org/10.1007/s00726-017-2493-3
|
| [156] |
Neis EPJG, Dejong CHC, Rensen SS (2015) The role of microbial amino acid metabolism in host metabolism. Nutrients 7: 2930-2946. https://doi.org/10.3390/nu7042930
|
| [157] |
Birklein F, Schmelz M (2008) Neuropeptides, neurogenic inflammation and complex regional pain syndrome (CRPS). Neurosci Lett 437: 199-202. https://doi.org/10.1016/j.neulet.2008.03.081
|
| [158] |
Seifert O, Baerwald C (2021) Interaction of pain and chronic inflammation. Z Rheumatol 80: 205-213. https://doi.org/10.1007/s00393-020-00951-8
|
| [159] |
Costigan M, Scholz J, Woolf CJ (2009) Neuropathic pain: a maladaptive response of the nervous system to damage. Annu Rev Neurosci 32: 1-32. https://doi.org/10.1146/annurev.neuro.051508.135531
|
| [160] |
Dinan TG, Cryan JF (2017) The Microbiome-Gut-Brain Axis in Health and Disease. Gastroenterol Clin North Am 46: 77-89. https://doi.org/10.1016/j.gtc.2016.09.007
|
| [161] |
Hu S, Png E, Gowans M, et al. (2021) Ectopic gut colonization: a metagenomic study of the oral and gut microbiome in Crohn's disease. Gut Pathog 13: 13. https://doi.org/10.1186/s13099-021-00409-5
|
| [162] |
Vinolo MAR, Rodrigues HG, Nachbar RT, et al. (2011) Regulation of inflammation by short chain fatty acids. Nutrients 3: 858-876. https://doi.org/10.3390/nu3100858
|
| [163] |
Tedelind S, Westberg F, Kjerrulf M, et al. (2007) Anti-inflammatory properties of the short-chain fatty acids acetate and propionate: a study with relevance to inflammatory bowel disease. World J Gastroenterol 13: 2826-2832. https://doi.org/10.3748/wjg.v13.i20.2826
|
| [164] |
Wang F, Liu J, Weng T, et al. (2017) The Inflammation Induced by Lipopolysaccharide can be Mitigated by Short-chain Fatty Acid, Butyrate, through Upregulation of IL-10 in Septic Shock. Scand J Immunol 85: 258-263. https://doi.org/10.1111/sji.12515
|
| [165] |
Handa AK, Fatima T, Mattoo AK (2018) Polyamines: Bio-Molecules with Diverse Functions in Plant and Human Health and Disease. Front Chem 6: 10. https://doi.org/10.3389/fchem.2018.00010
|
| [166] |
Robertson RC, Seira Oriach C, Murphy K, et al. (2017) Deficiency of essential dietary n-3 PUFA disrupts the caecal microbiome and metabolome in mice. Br J Nutr 118: 959-970. https://doi.org/10.1017/S0007114517002999
|
| [167] |
Goldberg RJ, Katz J (2007) A meta-analysis of the analgesic effects of omega-3 polyunsaturated fatty acid supplementation for inflammatory joint pain. Pain 129: 210-223. https://doi.org/10.1016/j.pain.2007.01.020
|
| [168] |
Spagnuolo C, Moccia S, Russo GL (2018) Anti-inflammatory effects of flavonoids in neurodegenerative disorders. Eur J Med Chem 153: 105-115. https://doi.org/10.1016/j.ejmech.2017.09.001
|
| [169] |
Cheng H, Zhang Y, Lu W, et al. (2018) Caffeic acid phenethyl ester attenuates neuropathic pain by suppressing the p38/NF-κB signal pathway in microglia. J Pain Res 11: 2709-2719. https://doi.org/10.2147/JPR.S166274
|
| [170] |
Filho AW, Filho VC, Olinger L, et al. (2008) Quercetin: further investigation of its antinociceptive properties and mechanisms of action. Arch Pharm Res 31: 713-721. https://doi.org/10.1007/s12272-001-1217-2
|
| [171] | Di Pierro F, Zacconi P, Bertuccioli A, et al. (2017) A naturally-inspired, curcumin-based lecithin formulation (Meriva® formulated as the finished product Algocur®) alleviates the osteo-muscular pain conditions in rugby players. Eur Rev Med Pharmacol Sci 21: 4935-4940. |
| [172] |
Chariot P, Bignani O (2003) Skeletal muscle disorders associated with selenium deficiency in humans. Muscle Nerve 27: 662-668. https://doi.org/10.1002/mus.10304
|
| [173] |
Castro J, Cooney MF (2017) Intravenous Magnesium in the Management of Postoperative Pain. J Perianesth Nurs 32: 72-76. https://doi.org/10.1016/j.jopan.2016.11.007
|
| [174] |
Gaikwad M, Vanlint S, Moseley GL, et al. (2018) Factors Associated with Vitamin D Testing, Deficiency, Intake, and Supplementation in Patients with Chronic Pain. J Diet Suppl 15: 636-648. https://doi.org/10.1080/19390211.2017.1375060
|
| [175] |
de Rienzo-Madero B, Coffeen U, Simón-Arceo K, et al. (2013) Taurine enhances antinociception produced by a COX-2 inhibitor in an inflammatory pain model. Inflammation 36: 658-664. https://doi.org/10.1007/s10753-012-9589-4
|
| [176] |
Ziółkowska B (2021) The Role of Mesostriatal Dopamine System and Corticostriatal Glutamatergic Transmission in Chronic Pain. Brain Sci 11: 1311. https://doi.org/10.3390/brainsci11101311
|
| [177] |
Heiss CN, Olofsson LE (2018) Gut Microbiota-Dependent Modulation of Energy Metabolism. J Innate Immun 10: 163-171. https://doi.org/10.1159/000481519
|
| [178] |
Lach G, Schellekens H, Dinan TG, et al. (2018) Anxiety, Depression, and the Microbiome: A Role for Gut Peptides. Neurotherapeutics 15: 36-59. https://doi.org/10.1007/s13311-017-0585-0
|
| [179] |
Liang Y, Ma Y, Wang J, et al. (2021) Leptin Contributes to Neuropathic Pain via Extrasynaptic NMDAR-nNOS Activation. Mol Neurobiol 58: 1185-1195. https://doi.org/10.1007/s12035-020-02180-1
|
| [180] |
Hannibal KE, Bishop MD (2014) Chronic stress, cortisol dysfunction, and pain: a psychoneuroendocrine rationale for stress management in pain rehabilitation. Phys Ther 94: 1816-1825. https://doi.org/10.2522/ptj.20130597
|
| [181] |
Lee DY, Kim E, Choi MH (2015) Technical and clinical aspects of cortisol as a biochemical marker of chronic stress. BMB Rep 48: 209-216. https://doi.org/10.5483/bmbrep.2015.48.4.275
|
| [182] |
Ley RE (2016) Gut microbiota in 2015: Prevotella in the gut: choose carefully. Nat Rev Gastroenterol Hepatol 13: 69-70. https://doi.org/10.1038/nrgastro.2016.4
|
| [183] |
Iacob S, Iacob DG (2019) Infectious Threats, the Intestinal Barrier, and Its Trojan Horse: Dysbiosis. Front Microbiol 10: 1676. https://doi.org/10.3389/fmicb.2019.01676
|
| [184] |
Schroeder BO (2019) Fight them or feed them: how the intestinal mucus layer manages the gut microbiota. Gastroenterol Rep (Oxf) 7: 3-12. https://doi.org/10.1093/gastro/goy052
|
| [185] |
Sun L, Yang S, Deng Q, et al. (2020) Salmonella Effector SpvB Disrupts Intestinal Epithelial Barrier Integrity for Bacterial Translocation. Front Cell Infect Microbiol 10: 606541. https://doi.org/10.3389/fcimb.2020.606541
|
| [186] |
Donati Zeppa S, Agostini D, Gervasi M, et al. (2020) Mutual Interactions among Exercise, Sport Supplements and Microbiota. Nutrients 12: 17. https://doi.org/10.3390/nu12010017
|
| [187] |
Lin Y, Zhang Y (2019) Renoprotective effect of oral rehydration solution III in exertional heatstroke rats. Ren Fail 41: 190-196. https://doi.org/10.1080/0886022X.2019.1590211
|
| [188] |
King MA, Leon LR, Mustico DL, et al. (2015) Biomarkers of multiorgan injury in a preclinical model of exertional heat stroke. J Appl Physiol 118: 1207-1220. https://doi.org/10.1152/japplphysiol.01051.2014
|
| [189] |
Wacker C, Prkno A, Brunkhorst FM, et al. (2013) Procalcitonin as a diagnostic marker for sepsis: a systematic review and meta-analysis. Lancet Infect Dis 13: 426-435. https://doi.org/10.1016/S1473-3099(13)70301-3
|
| [190] | Brock-Utne JG, Gaffin SL, Wells MT, et al. (1988) Endotoxaemia in exhausted runners after a long-distance race. S Afr Med J 73: 533-536. |
| [191] |
Cook MD, Allen JM, Pence BD, et al. (2016) Exercise and gut immune function: evidence of alterations in colon immune cell homeostasis and microbiome characteristics with exercise training. Immunol Cell Biol 94: 158-163. https://doi.org/10.1038/icb.2015.108
|
| [192] | Spagnuolo PA, Hoffman-Goetz L (2009) Effect of dextran sulfate sodium and acute exercise on mouse intestinal inflammation and lymphocyte cytochrome c levels. J Sports Med Phys Fitness 49: 112-121. |
| [193] |
Shandilya S, Kumar S, Kumar Jha N, et al. (2021) Interplay of gut microbiota and oxidative stress: Perspective on neurodegeneration and neuroprotection. J Adv Res 38: 223-244. https://doi.org/10.1016/j.jare.2021.09.005
|
| [194] |
Karl JP, Berryman CE, Young AJ, et al. (2018) Associations between the gut microbiota and host responses to high altitude. Am J Physiol Gastrointest Liver Physiol 315: G1003-G1015. https://doi.org/10.1152/ajpgi.00253.2018
|
| [195] |
Clarke SF, Murphy EF, O'Sullivan O, et al. (2014) Exercise and associated dietary extremes impact on gut microbial diversity. Gut 63: 1913-1920. http://doi.org/10.1136/gutjnl-2013-306541
|
| [196] |
Mittinty MM, Lee JY, Walton DM, et al. (2022) Integrating the Gut Microbiome and Stress-Diathesis to Explore Post-Trauma Recovery: An Updated Model. Pathogens 11: 716. https://doi.org/10.3390/pathogens11070716
|
| [197] |
David LA, Maurice CF, Carmody RN, et al. (2014) Diet rapidly and reproducibly alters the human gut microbiome. Nature 505: 559-563. https://doi.org/10.1038/nature12820
|
| [198] |
Jernberg C, Löfmark S, Edlund C, et al. (2010) Long-term impacts of antibiotic exposure on the human intestinal microbiota. Microbiology (Reading) 156: 3216-3223. https://doi.org/10.1099/mic.0.040618-0
|
| [199] | Barton W, Penney NC, Cronin O, et al. (2018) The microbiome of professional athletes differs from that of more sedentary subjects in composition and particularly at the functional metabolic level. Gut 67: 625-633. https://doi.org/10.1136/gutjnl-2016-313627 |
| [200] |
Oda H (2015) Chrononutrition. J Nutr Sci Vitaminol (Tokyo) 61 Suppl: S92-S94. https://doi.org/10.3177/jnsv.61.S92
|


Figures(3)

Jesús Álvarez-Herms, Adriana González, Francisco Corbi, Iñaki Odriozola, Adrian Odriozola. Possible relationship between the gut leaky syndrome and musculoskeletal injuries: the important role of gut microbiota as indirect modulator[J]. AIMS Public Health, 2023, 10(3): 710-738. doi: 10.3934/publichealth.2023049







 DownLoad:
DownLoad: