Citation: Viktor Semenkov, Anatolii Mikhalskii, Alexandr Sapoznikov. Influence of heat shock proteins in individual sensitivity of human neutrophils to heat stress[J]. AIMS Molecular Science, 2019, 6(2): 38-51. doi: 10.3934/molsci.2019.2.38

| [1] |
Wang J, Grishin AV, Ford HR (2016) Experimental anti-inflammatory drug semapimod inhibits Toll-like receptor signaling by targeting the TLR chaperone gp96. J Immunol 196: 5130–5137. doi: 10.4049/jimmunol.1502135

|
| [2] | Gao H, Liu X, Sun W, et al. (2017) Total transhinones exhibits anti-inflammatory effects through bloking TLR4 dimerization via the MyD88 pathway. Cell Death Dis 8: e3004. |
| [3] |
Speer EM, Dowling DJ, Xu J, et al. (2018) Pentoxifylline, dexamethasone and azithromycin demonstrate distinct age-dependent and synergistic inhibition of TLR- and inflammasome-mediated cytokine production in human newborn and adult blood in vitro. PLoS ONE 13: e0196352. doi: 10.1371/journal.pone.0196352
|
| [4] |
Ikwegbue PC, Masamba P, Oyinloye BE, et al. (2018) Roles of heat shock proteins in apoptosis, oxidative stress, human inflammatory diseases, and cancer. Pharmaceuticals 11: 2. doi: 10.3390/pharmaceutics11010002
|
| [5] |
Uttara B, Singh AV, Zamboni P, et al. (2009) Oxidative stress and neurodegenerative diseases: A review of upstream and downstream antioxidant therapeutic options. Curr Neuropharmacol 7: 65–74. doi: 10.2174/157015909787602823
|
| [6] |
Maridonneau-Parini I, Clerc J, Polla BS (1988) Heat shock inhibits NADPH oxidase in human neutrophils. Biochem Biophys Res Commun 154: 179–186. doi: 10.1016/0006-291X(88)90667-5
|
| [7] | Costa VM, Silva R, Ferreira LM, et al. (2007) Oxidation process of adrenaline in freshly isolated rat cardiomyocytes: Formation of adrenochrome, quinoproteins, and GSH adduct. Chem Res Toxicol 8: 1183–1191. |
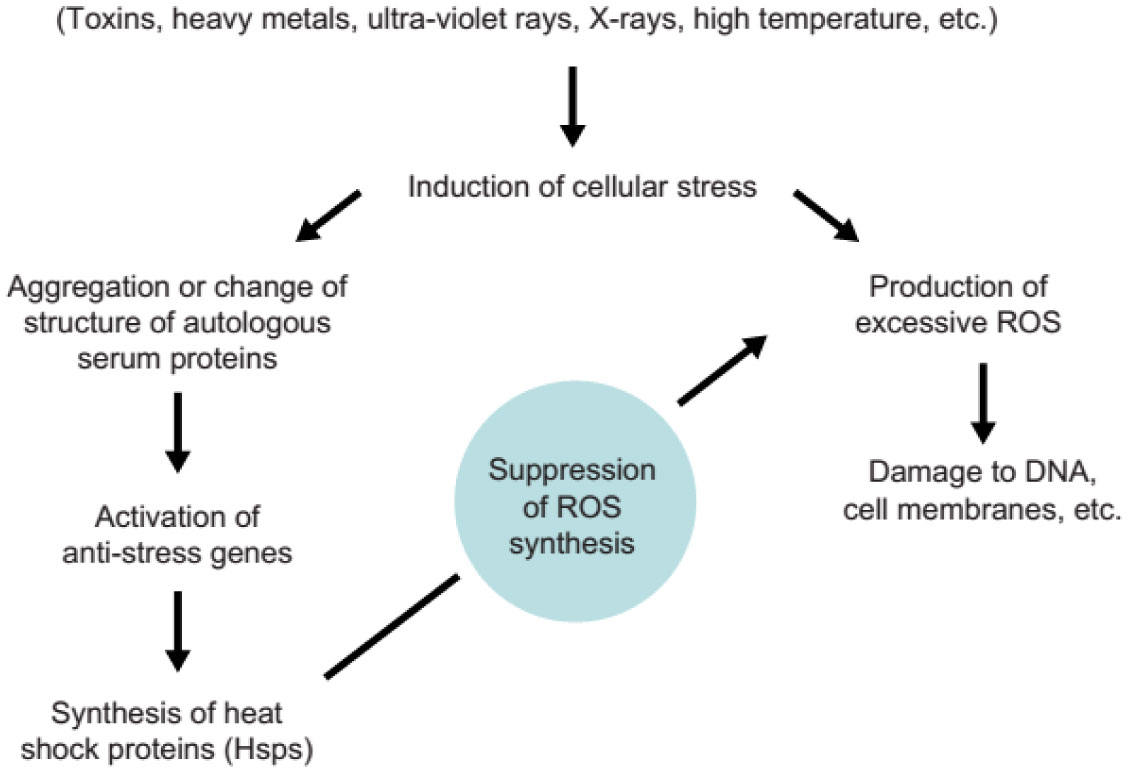
| [8] | Kovalenko EI, Boyko AA, Semenkov VF, et al. (2014) ROS production, intracellular HSP70 levels and their relationship in human neutrophils: Effects of age. Oncotarget 5: 11800–11812. |
| [9] |
Allen RC, Loose LD (1976) Phagocytic activation of a luminoldependent chemiluminescence in rabbit alveolar and peritoneal macrophages. Biochem Biophys Res Commun 69: 245–252. doi: 10.1016/S0006-291X(76)80299-9
|
| [10] |
Doyle SE, O'Connell RM, Miranda GA, et al. (2004) Toll-like receptors induce a phagocytic gene program through p38. J Exp Med 199: 81–90. doi: 10.1084/jem.20031237
|
| [11] |
Hermann JK, Lin S, Soffer A, et al. (2018) The role of Toll-like receptor 2 and 4 innate immunity pathways in intracortical microelectrode-induced neuroinflammation. Front Bioeng Biotechnol 6: 113. doi: 10.3389/fbioe.2018.00113
|
| [12] |
Ogava K, Suzuki K, Okutsu M, et al. (2008) The association of elevated reactive oxygen levels from neutrophils with low-grade inflammation in the elderly. Immun Ageing 5: 13–20. doi: 10.1186/1742-4933-5-13
|
| [13] |
Singh R, Kolvraa S, Rattan SIS (2007) Genetics of human longevity with emphasis on the relevance of HSP70 as candidate genes. Front Biosci 12: 4504–4513. doi: 10.2741/2405
|
| [14] | Semenkov VF, Michalski AI, Sapozhnikov AM (2015) Heating and ultraviolet light activate anti-stress gene functions in humans. Front Genet 6: 245. |
| [15] |
Chittiboyina S, Bai Y, Lelievre SA (2018) Microenvironment-cell nucleus relationship in the context of oxidative stress. Front Cell Dev Biol 6: 23. doi: 10.3389/fcell.2018.00023
|
| [16] |
Zininga T, Ramatsui L, Shonhai A (2018) Heat shock proteins as immunomodulants. Molecules 23: 2846. doi: 10.3390/molecules23112846
|
| [17] | Guisasola MC, Alonso B, Bravo B, et al. (2017) An overview of cytokines and heat shock response in polytraumatized patients. Cell Stress Chaperones 23: 483–489. |
Figures(10) / Tables(6)

Viktor Semenkov, Anatolii Mikhalskii, Alexandr Sapoznikov. Influence of heat shock proteins in individual sensitivity of human neutrophils to heat stress[J]. AIMS Molecular Science, 2019, 6(2): 38-51. doi: 10.3934/molsci.2019.2.38







 DownLoad:
DownLoad: