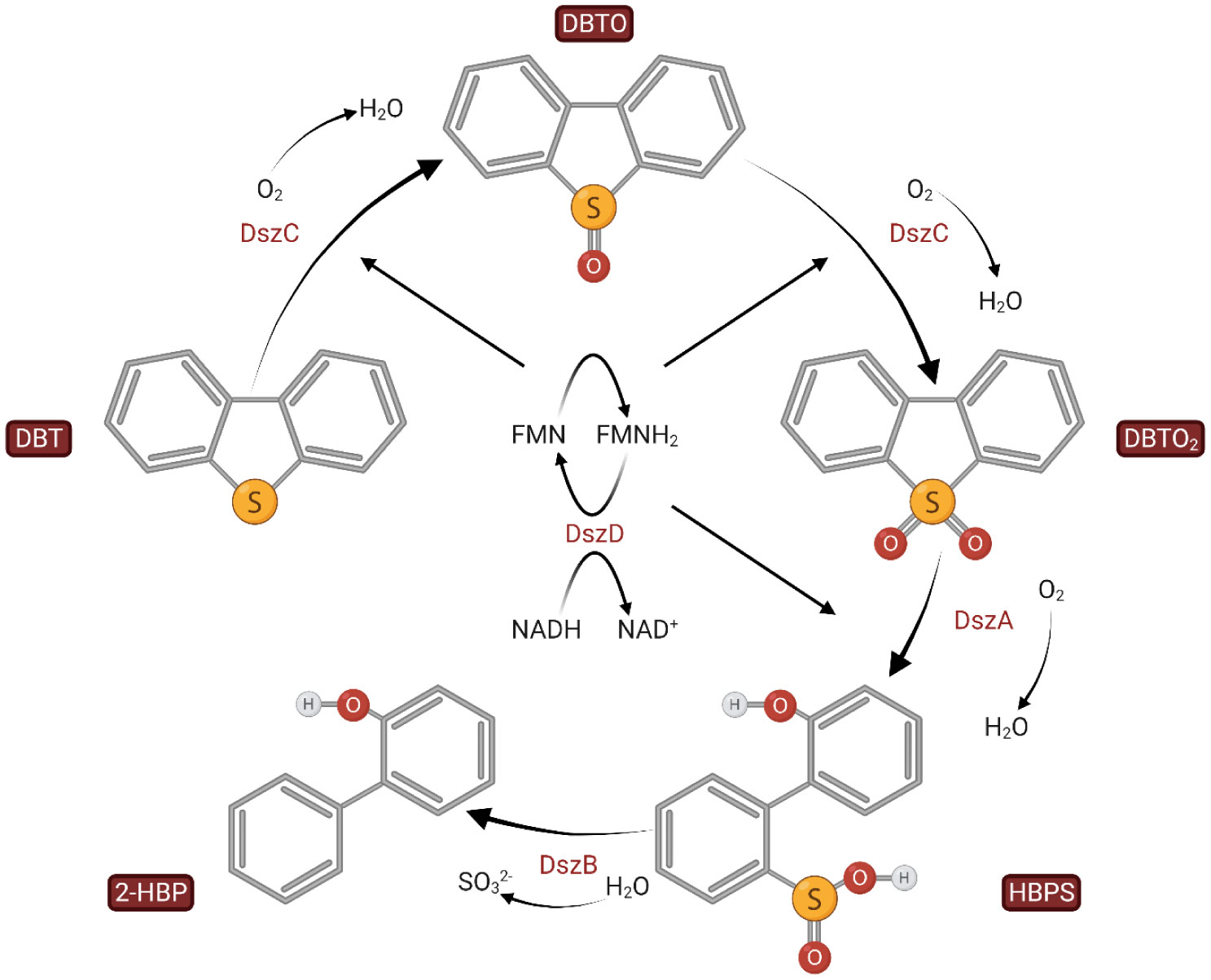
Sustainable biodesulfurization (BDS) processes require the use of microbial biocatalysts that display high activity against the recalcitrant heterocyclic sulfur compounds and can simultaneously withstand the harsh conditions of contact with petroleum products, inherent to any industrial biphasic BDS system. In this framework, the functional microbial BDS-related diversity in a naturally oil-exposed ecosystem, was examined through a 4,6-dimethyl-dibenzothiophene based enrichment process. Two new Rhodococcus sp. strains were isolated, which during a medium optimization process revealed a significantly enhanced BDS activity profile when compared to the model strain R. qingshengii IGTS8. In biocatalyst stability studies conducted in biphasic mode using partially hydrodesulfurized diesel under various process conditions, the new strains also presented an enhanced stability phenotype. In these studies, it was also demonstrated for all strains, that the BDS activity losses were decoupled from the overall cells' viability, in addition to the fact that the use of whole-broth biocatalyst positively affected BDS performance.
Citation: Panayiotis D. Glekas, Olga Martzoukou, Maria-Eleni Mastrodima, Efstathios Zarkadoulas, Dimitrios S. Kanakoglou, Dimitris Kekos, Michalis Pachnos, George Mavridis, Diomi Mamma, Dimitris G. Hatzinikolaou. Deciphering the biodesulfurization potential of two novel Rhodococcus isolates from a unique Greek environment[J]. AIMS Microbiology, 2022, 8(4): 484-506. doi: 10.3934/microbiol.2022032
Sustainable biodesulfurization (BDS) processes require the use of microbial biocatalysts that display high activity against the recalcitrant heterocyclic sulfur compounds and can simultaneously withstand the harsh conditions of contact with petroleum products, inherent to any industrial biphasic BDS system. In this framework, the functional microbial BDS-related diversity in a naturally oil-exposed ecosystem, was examined through a 4,6-dimethyl-dibenzothiophene based enrichment process. Two new Rhodococcus sp. strains were isolated, which during a medium optimization process revealed a significantly enhanced BDS activity profile when compared to the model strain R. qingshengii IGTS8. In biocatalyst stability studies conducted in biphasic mode using partially hydrodesulfurized diesel under various process conditions, the new strains also presented an enhanced stability phenotype. In these studies, it was also demonstrated for all strains, that the BDS activity losses were decoupled from the overall cells' viability, in addition to the fact that the use of whole-broth biocatalyst positively affected BDS performance.

| [1] |
Mohebali G, Ball AS (2016) Biodesulfurization of diesel fuels–Past, present and future perspectives. Int Biodeterior Biodegradation 110: 163-180. https://doi.org/10.1016/j.ibiod.2016.03.011 
|
| [2] |
Kilbane JJ (2017) Biodesulfurization: How to make it work?. Arab J Sci Eng 42: 1-9. https://doi.org/10.1007/s13369-016-2269-1
|
| [3] |
Wang J, Butler RR, Wu F, et al. (2017) Enhancement of microbial biodesulfurization via genetic engineering and adaptive evolution. PLoS One 12: e0168833. https://doi.org/10.1371/journal.pone.0168833
|
| [4] | Xu P, Feng J, Yu B, et al. (2009) Recent developments in biodesulfurization of fossil fuels. Adv Biochem Eng Biotechnol 113: 255-274. https://doi.org/10.1007/10_2008_16 |
| [5] |
Boniek D, Figueiredo D, dos Santos AFB, et al. (2015) Biodesulfurization: a mini review about the immediate search for the future technology. Clean Technol Environ Policy 17: 29-37. https://doi.org/10.1007/s10098-014-0812-x
|
| [6] |
Bordoloi NK, Rai SK, Chaudhuri MK, et al. (2014) Deep-desulfurization of dibenzothiophene and its derivatives present in diesel oil by a newly isolated bacterium Achromobacter sp. to reduce the environmental pollution from fossil fuel combustion. Fuel Process Technol 119: 236-244. https://doi.org/10.1016/j.fuproc.2013.10.014
|
| [7] |
Okada H, Nomura N, Nakahara T, et al. (2002) Analysis of dibenzothiophene metabolic pathway in Mycobacterium strain G3. J Biosci Bioeng 93: 491-497. https://doi.org/10.1016/S1389-1723(02)80097-4
|
| [8] |
Chen H, Cai YB, Zhang WJ, et al. (2009) Methoxylation pathway in biodesulfurization of model organosulfur compounds with Mycobacterium sp. Bioresour Technol 100: 2085-2087. https://doi.org/10.1016/j.biortech.2008.10.010
|
| [9] |
Gallagher JR, Olson ES, Stanley DC (1993) Microbial desulfurization of dibenzothiophene: A sulfur-specific pathway. FEMS Microbiol Lett 107: 31-35. https://doi.org/10.1111/j.1574-6968.1993.tb05999.x
|
| [10] |
Thompson D, Cognat V, Goodfellow M, et al. (2020) Phylogenomic classification and biosynthetic potential of the fossil fuel-biodesulfurizing Rhodococcus strain IGTS8. Front Microbiol 11: 1417. https://doi.org/10.3389/fmicb.2020.01417
|
| [11] |
Martinez I, Santos VE, Alcon A, et al. (2015) Enhancement of the biodesulfurization capacity of Pseudomonas putida CECT5279 by co-substrate addition. Process Biochem 50: 119-124. https://doi.org/10.1016/j.procbio.2014.11.001
|
| [12] |
Mawad AMM, Hassanein M, Aldaby ES, et al. (2021) Desulphurisation kinetics of thiophenic compound by sulphur oxidizing Klebsiella oxytoca SOB-1. J Appl Microbiol 130: 1181-1191. https://doi.org/10.1111/jam.14829
|
| [13] | Mohamed MES, Al-Yacoub ZH, Vedakumar JV (2015) Biocatalytic desulfurization of thiophenic compounds and crude oil by newly isolated bacteria. Front Microbiol 6: 112. https://doi.org/10.3389/fmicb.2015.00112 |
| [14] |
Bhanjadeo MM, Rath K, Gupta D, et al. (2018) Differential desulfurization of dibenzothiophene by newly identified MTCC strains: Influence of Operon Array. PLoS One 13: e0192536. https://doi.org/10.1371/journal.pone.0192536
|
| [15] |
Martínez I, Mohamed ME-S, Rozas D, et al. (2016) Engineering synthetic bacterial consortia for enhanced desulfurization and revalorization of oil sulfur compounds. Metab Eng 35: 46-54. https://doi.org/10.1016/j.ymben.2016.01.005
|
| [16] |
Kilbane JJ (2006) Microbial biocatalyst developments to upgrade fossil fuels. Curr Opin Biotechnol 17: 305-314. https://doi.org/10.1016/j.copbio.2006.04.005
|
| [17] |
Schade T, Andersson JT (2006) Speciation of alkylated dibenzothiophenes in a deeply desulfurized diesel fuel. Energy Fuels 20: 1614-1620. https://doi.org/10.1021/ef0502507
|
| [18] |
Chen S, Zhao C, Liu Q, et al. (2019) Biodesulfurization of diesel oil in oil-water two phase reaction system by Gordonia sp. SC-10. Biotechnol Lett 41: 547-554. https://doi.org/10.1007/s10529-019-02663-9
|
| [19] |
Mingfang L, Jianmin X, Zhongxuan G, et al. (2003) Microbial desulfurization of dibenzothiophene and 4,6-dimethyldibenzothiophene in dodecane and straight-run diesel oil. Korean J Chem Eng 20: 702-704. https://doi.org/10.1007/BF02706911
|
| [20] |
Awadh M, Mahmoud H, Abed RMM, et al. (2020) Diesel-born organosulfur compounds stimulate community re-structuring in a diesel-biodesulfurizing consortium. Biotechnol Rep (Amst) 28: e00572. https://doi.org/10.1016/j.btre.2020.e00572
|
| [21] |
Avramidis P, Kalaitzidis S, Iliopoulos G, et al. (2017) The so called ‘Herodotus Springs’ at ‘Keri Lake’ in Zakynthos Island western Greece: A palaeoenvironmental and palaeoecological approach. Quat Int 439: 37-51. https://doi.org/10.1016/j.quaint.2016.12.014
|
| [22] |
Pasadakis N, Dagounaki V, Chamilaki E (2016) A comparative organic geochemical study of oils seeps in Western Greece. Energy Sources Part A 38: 362-369. https://doi.org/10.1080/15567036.2013.766660
|
| [23] | Tamura K, Nei M (1993) Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol Biol Evol 10: 512-526. |
| [24] |
Gascuel O (1997) BIONJ: an improved version of the NJ algorithm based on a simple model of sequence data. Mol Biol Evol 14: 685-695. https://doi.org/10.1093/oxfordjournals.molbev.a025808
|
| [25] |
Tamura K, Stecher G, Kumar S (2021) MEGA11: Molecular evolutionary genetics analysis version 11. Mol Biol Evol 38: 3022-3027. https://doi.org/10.1093/molbev/msab120
|
| [26] |
Letunic I, Bork P (2021) Interactive Tree Of Life (iTOL) v5: an online tool for phylogenetic tree display and annotation. Nucleic Acids Res 49: W293-W296. https://doi.org/10.1093/nar/gkab301
|
| [27] |
Martzoukou O, Glekas PD, Avgeris M, et al. (2022) Interplay between sulfur assimilation and biodesulfurization activity in Rhodococcus qingshengii IGTS8: Insights into a regulatory role of the reverse transsulfuration pathway. mBio 13: e0075422. https://doi.org/10.1128/mbio.00754-22
|
| [28] |
Prasoulas G, Dimos K, Glekas P, et al. (2021) Biodesulfurization of dibenzothiophene and its alkylated derivatives in a two-phase bubble column bioreactor by resting cells of Rhodococcus erythropolis IGTS8. Processes 9: 2064. https://doi.org/10.3390/pr9112064
|
| [29] |
Olmo CH del, Santos VE, Alcon A, et al. (2005) Production of a Rhodococcus erythropolis IGTS8 biocatalyst for DBT biodesulfurization: influence of operational conditions. Biochem Eng J 22: 229-237. https://doi.org/10.1016/j.bej.2004.09.015
|
| [30] |
Hirschler A, Carapito C, Maurer L, et al. (2021) Biodesulfurization induces reprogramming of sulfur metabolism in Rhodococcus qingshengii IGTS8: Proteomics and untargeted metabolomics. Microbiol Spectr : e0069221. https://doi.org/10.1128/Spectrum.00692-21
|
| [31] |
Murarka P, Bagga T, Singh P, et al. (2019) Isolation and identification of a TetR family protein that regulates the biodesulfurization operon. AMB Express 9: 71. https://doi.org/10.1186/s13568-019-0801-x
|
| [32] | Ismail W, El-Sayed WS, Abdul Raheem AS, et al. (2016) Biocatalytic desulfurization capabilities of a mixed culture during non-destructive utilization of recalcitrant organosulfur compounds. Front Microbiol 7: 266. https://doi.org/10.3389/fmicb.2016.00266 |
| [33] |
Akhtar N, Ghauri MA, Akhtar K (2016) Dibenzothiophene desulfurization capability and evolutionary divergence of newly isolated bacteria. Arch Microbiol 198: 509-519. https://doi.org/10.1007/s00203-016-1209-5
|
| [34] |
Peng C, Huang D, Shi Y, et al. (2019) Comparative transcriptomic analysis revealed the key pathways responsible for organic sulfur removal by thermophilic bacterium Geobacillus thermoglucosidasius W-2. Sci Total Environ 676: 639-650. https://doi.org/10.1016/j.scitotenv.2019.04.328
|
| [35] |
Khedkar S, Shanker R (2014) Degradation of dibenzothiophene and its metabolite 3-hydroxy-2-formylbenzothiophene by an environmental isolate. Biodegradation 25: 643-654. https://doi.org/10.1007/s10532-014-9688-z
|
| [36] |
Wang L, Ji G, Huang S (2019) Contribution of the Kodama and 4S pathways to the dibenzothiophene biodegradation in different coastal wetlands under different C/N ratios. J Environ Sci 76: 217-226. https://doi.org/10.1016/j.jes.2018.04.029
|
| [37] |
Piccoli S, Andreolli M, Giorgetti A, et al. (2014) Identification of aldolase and ferredoxin reductase within the dbt operon of Burkholderia fungorum DBT1. J Basic Microbiol 54: 464-469. https://doi.org/10.1002/jobm.201200408
|
| [38] |
Li L, Shen X, Zhao C, et al. (2019) Biodegradation of dibenzothiophene by efficient Pseudomonas sp. LKY-5 with the production of a biosurfactant. Ecotoxicol Environ Saf 176: 50-57. https://doi.org/10.1016/j.ecoenv.2019.03.070
|
| [39] |
Wang W, Ma T, Lian K, et al. (2013) Genetic analysis of benzothiophene biodesulfurization pathway of Gordonia terrae strain C-6. PLoS One 8: e84386. https://doi.org/10.1371/journal.pone.0084386
|
| [40] |
del Olmo CH, Alcon A, Santos VE, et al. (2005) Modeling the production of a Rhodococcus erythropolis IGTS8 biocatalyst for DBT biodesulfurization: Influence of media composition. Enzyme Microb Technol 37: 157-166. https://doi.org/10.1016/j.enzmictec.2004.06.016
|
| [41] |
Teixeira AV, Paixão SM, da Silva TL, et al. (2014) Influence of the carbon source on Gordonia alkanivorans strain 1B resistance to 2-hydroxybiphenyl toxicity. Appl Biochem Biotechnol 173: 870-882. https://doi.org/10.1007/s12010-014-0902-6
|
| [42] |
Li MZ, Squires CH, Monticello DJ, et al. (1996) Genetic analysis of the dsz promoter and associated regulatory regions of Rhodococcus erythropolis IGTS8. J Bacteriol 178: 6409-6418. https://doi.org/10.1128/jb.178.22.6409-6418.1996
|
| [43] |
Hai Y, Kishimoto M, Omasa T, et al. (2000) Increase in desulfurization activity of Rhodococcus erythropolis KA2-5-l using ethanol feeding. J Biosci Bioeng 89: 361-366. https://doi.org/10.1016/S1389-1723(00)88959-8
|
| [44] |
Dejaloud A, Habibi A, Vahabzadeh F (2020) DBT desulfurization by Rhodococcus erythropolis PTCC 1767 in aqueous and biphasic systems. Chem Pap 74: 3605-3615. https://doi.org/10.1007/s11696-020-01191-5
|
| [45] |
Nassar HN, Deriase SF, El-Gendy NS (2017) Statistical optimization of biomass production and biodesulfurization activity of Rhodococcus erythropolis HN2. Pet Sci Technol 35: 1951-1959. https://doi.org/10.1080/10916466.2017.1373129
|
| [46] |
Konishi M, Kishimoto M, Omasa T, et al. (2005) Effect of sulfur sources on specific desulfurization activity of Rhodococcus erythropolis KA2-5-1 in exponential fed-batch culture. J Biosci Bioeng 99: 259-263. https://doi.org/10.1263/jbb.99.259
|
| [47] |
Aggarwal S, Karimi IA, Lee DY (2011) Flux-based analysis of sulfur metabolism in desulfurizing strains of Rhodococcus erythropolis. FEMS Microbiol Lett 315: 115-121. https://doi.org/10.1111/j.1574-6968.2010.02179.x
|
| [48] |
Silva TP, Alves L, Paixão SM (2020) Effect of dibenzothiophene and its alkylated derivatives on coupled desulfurization and carotenoid production by Gordonia alkanivorans strain 1B. J Environ Manage 270: 110825. https://doi.org/10.1016/j.jenvman.2020.110825
|
| [49] |
Tanaka Y, Yoshikawa O, Maruhashi K, et al. (2002) The cbs mutant strain of Rhodococcus erythropolis KA2-5-1 expresses high levels of Dsz enzymes in the presence of sulfate. Arch Microbiol 178: 351-357. https://doi.org/10.1007/s00203-002-0466-7
|
| [50] |
Mohebali G, Ball AS, Kaytash A, et al. (2008) Dimethyl sulfoxide (DMSO) as the sulfur source for the production of desulfurizing resting cells of Gordonia alkanivorans RIPI90A. Microbiology 154: 878-885. https://doi.org/10.1099/mic.0.2007/013011-0
|
| [51] |
Abbad-Andaloussi S, Lagnel C, Warzywoda M, et al. (2003) Multi-criteria comparison of resting cell activities of bacterial strains selected for biodesulfurization of petroleum compounds. Enzyme Microb Technol 32: 446-454. https://doi.org/10.1016/S0141-0229(02)00320-4
|
| [52] |
Blasco R, Martínez-Luque M, Madrid MP, et al. (2001) Rhodococcus sp. RB1 grows in the presence of high nitrate and nitrite concentrations and assimilates nitrate in moderately saline environments. Arch Microbiol 175: 435-440. https://doi.org/10.1007/s002030100285
|
| [53] |
Pacheco GJ, Ciapina EMP, Gomes E de B, et al. (2010) Biosurfactant production by Rhodococcus erythropolis and its application to oil removal. Braz J Microbiol 41: 685-693. https://doi.org/10.1590/S1517-83822010000300019
|
| [54] |
Styczynski M, Rogowska A, Gieczewska K, et al. (2020) Genome-based insights into the production of carotenoids by antarctic bacteria, Planococcus sp. ANT_H30 and Rhodococcus sp. ANT_H53B. Molecules 25: 4375. https://doi.org/10.3390/molecules25194357
|
| [55] |
Michas A, Vestergaard G, Trautwein K, et al. (2017) More than 2500 years of oil exposure shape sediment microbiomes with the potential for syntrophic degradation of hydrocarbons linked to methanogenesis. Microbiome 5: 118. https://doi.org/10.1186/s40168-017-0337-8
|
| [56] |
Carr GJ, Ferguson SJ (1990) Nitric oxide formed by nitrite reductase of Paracoccus denitrificans is sufficiently stable to inhibit cytochrome oxidase activity and is reduced by its reductase under aerobic conditions. Biochim Biophys Acta 1017: 57-62. https://doi.org/10.1016/0005-2728(90)90178-7
|
| [57] |
Maghsoudi S, Vossoughi M, Kheirolomoom A, et al. (2001) Biodesulfurization of hydrocarbons and diesel fuels by Rhodococcus sp. strain P32C1. Biochem Eng J 8: 151-156. https://doi.org/10.1016/S1369-703X(01)00097-3
|
| [58] |
Maass D, de Oliveira D, de Souza AAU, et al. (2014) Biodesulfurization of a system containing synthetic fuel using Rhodococcus erythropolis ATCC 4277. Appl Biochem Biotechnol 174: 2079-2085. https://doi.org/10.1007/s12010-014-1189-3
|
| [59] |
Hokmabadi M, Khosravinia S, Mahdavi MA, et al. (2022) Enhancing the biodesulphurization capacity of Rhodococcus sp. FUM94 in a biphasic system through optimization of operational factors. J Appl Microbiol 132: 3461-3475. https://doi.org/10.1111/jam.15442
|
| [60] |
Yang J, Hu Y, Zhao D, et al. (2007) Two-layer continuous-process design for the biodesulfurization of diesel oils under bacterial growth conditions. Biochem Eng J 37: 212-218. https://doi.org/10.1016/j.bej.2007.04.012
|
| [61] |
Yu B, Xu P, Shi Q, et al. (2006) Deep desulfurization of diesel oil and crude oils by a newly isolated Rhodococcus erythropolis strain. Appl Environ Microbiol 72: 54-58. https://doi.org/10.1128/AEM.72.1.54-58.2006
|
 microbiol-08-04-032-s001.pdf microbiol-08-04-032-s001.pdf |
 |





Figures(6) / Tables(1)

Panayiotis D. Glekas, Olga Martzoukou, Maria-Eleni Mastrodima, Efstathios Zarkadoulas, Dimitrios S. Kanakoglou, Dimitris Kekos, Michalis Pachnos, George Mavridis, Diomi Mamma, Dimitris G. Hatzinikolaou. Deciphering the biodesulfurization potential of two novel Rhodococcus isolates from a unique Greek environment[J]. AIMS Microbiology, 2022, 8(4): 484-506. doi: 10.3934/microbiol.2022032







 DownLoad:
DownLoad: