Citation: Luciana Porto de Souza Vandenberghe, Lina Marcela Blandon Garcia, Cristine Rodrigues, Marcela Cândido Camara, Gilberto Vinícius de Melo Pereira, Juliana de Oliveira, Carlos Ricardo Soccol. Potential applications of plant probiotic microorganisms in agriculture and forestry[J]. AIMS Microbiology, 2017, 3(3): 629-648. doi: 10.3934/microbiol.2017.3.629

| [1] |
Picard C, Baruffa E, Bosco M (2008) Enrichment and diversity of plant-probiotic microorganisms in the rhizosphere of hybrid maize during four growth cycles. Soil Biol Biochem 40: 106–115. doi: 10.1016/j.soilbio.2007.07.011

|
| [2] |
Benrebah F, Prevost D, Yezza A, et al. (2007) Agro-industrial waste materials and wastewater sludge for rhizobial inoculant production: A review. Bioresour Technol 98: 3535–3546. doi: 10.1016/j.biortech.2006.11.066
|
| [3] |
Mayak S, Tirosh T, Glick BR (2004) Plant growth-promoting bacteria confer resistance in tomato plants to salt stress. Plant Physiol Biochem 42: 565–572. doi: 10.1016/j.plaphy.2004.05.009
|
| [4] |
Singh JS, Pandey VC, Singh DP (2011) Efficient soil microorganisms: A new dimension for sustainable agriculture and environmental development. Agric Ecosyst Environ 140: 339–353. doi: 10.1016/j.agee.2011.01.017
|
| [5] | Gomez CG, Valero NV, Brigard RC (2012) Halotolerant/alkalophilic bacteria associated with the cyanobacterium Arthrospira platensis (Nordstedt) Gomont that promote early growth in Sorghum bicolor (L.). Moench Agron Colomb 30: 111–115. |
| [6] |
Yang J, Kloepper JW, Ryu CM (2009) Rhizosphere bacteria help plants tolerate abiotic stress. Trends Plant Sci 14: 1–4. doi: 10.1016/j.tplants.2008.10.004
|
| [7] |
Bhattacharyya D, Yu SM, Lee YH (2015) Volatile compounds from Alcaligenes faecalis JBCS1294 confer salt tolerance in Arabidopsis thaliana through the auxin and gibberellin pathways and differential modulation of gene expression in root and shoot tissues. Plant Growth Regul 75: 297–306. doi: 10.1007/s10725-014-9953-5
|
| [8] | Zhang Z, Lin W, Yang Y, et al. (2011) Effects of consecutively monocultured Rehmannia glutinosa L. on diversity of fungal community in rhizospheric soil. Agric Sci China 10: 1374–1384. |
| [9] | Zhang H, Kim MS, Sun Y, et al. (2008) Soil bacteria confer plant salt tolerance by tissue-specific regulation of the sodium transporter HKT1. Mol Plant Microbe In 21: 737–744. |
| [10] |
Porcel R, Zamarreño Á, García-Mina J, et al. (2014) Involvement of plant endogenous ABA in Bacillus megaterium PGPR activity in tomato plants. BMC Plant Biol 14: 36. doi: 10.1186/1471-2229-14-36
|
| [11] |
Tsuda K, Tsuji G, Higashiyama M, et al. (2016) Biological control of bacterial soft rot in Chinese cabbage by Lactobacillus plantarum strain BY under field conditions. Biol Control 100: 63–69. doi: 10.1016/j.biocontrol.2016.05.010
|
| [12] |
Waqas M, Khan AL, Kamran M, et al. (2012) Endophytic fungi produce gibberellins and indoleacetic acid and promotes host-plant growth during stress. Molecules 17: 10754–10773. doi: 10.3390/molecules170910754
|
| [13] | Khan AL, Hamayun M, Ahmad N, et al. (2011) Exophiala sp. LHL08 reprograms Cucumis sativus to higher growth under abiotic stresses. Physiol Plant 143: 329–343. |
| [14] |
Khan AL, Hamayun M, Ahmad N, et al. (2011) Salinity stress resistance offered by endophytic fungal interaction between Penicillium minioluteum LHL09 and glycine max. L. J Microbiol Biotechnol 21: 893–902. doi: 10.4014/jmb.1103.03012
|
| [15] | De Palma M, D'Agostino N, Proietti S, et al. (2016) Suppression Subtractive Hybridization analysis provides new insights into the tomato (Solanum lycopersicum L.) response to the plant probiotic microorganism Trichoderma longibrachiatum MK1. J Plant Physiol 190: 79–94. |
| [16] |
Qin Y, Druzhinina IS, Pan X, et al. (2016) Microbially mediated plant salt tolerance and microbiome-based solutions for saline agriculture. Biotechnol Adv 34: 1245–1259. doi: 10.1016/j.biotechadv.2016.08.005
|
| [17] |
Upadhyay SK, Singh JS, Saxena AK, et al. (2012) Impact of PGPR inoculation on growth and antioxidant status of wheat under saline conditions. Plant Biol 14: 605–611. doi: 10.1111/j.1438-8677.2011.00533.x
|
| [18] |
Redman RS, Kim YO, Woodward CJDA, et al. (2011) Increased fitness of rice plants to abiotic stress via habitat adapted symbiosis: A strategy for mitigating impacts of climate change. PLoS One 6: e14823. doi: 10.1371/journal.pone.0014823
|
| [19] |
Belimov AA, Dodd IC, Safronova VI, et al. (2014) Abscisic acid metabolizing rhizobacteria decrease ABA concentrations in planta and alter plant growth. Plant Physiol Biochem 74: 84–91. doi: 10.1016/j.plaphy.2013.10.032
|
| [20] |
Banaei-Asl F, Bandehagh A, Uliaei ED, et al. (2015) Proteomic analysis of canola root inoculated with bacteria under salt stress. J Proteomics 124: 88–111. doi: 10.1016/j.jprot.2015.04.009
|
| [21] | Melnick RL, Zidack NK, Bailey BA, et al. (2008) Bacterial endophytes: Bacillus spp. from annual crops as potential biological control agents of black pod rot of cacao. Biological Control 46: 46–56. |
| [22] |
Strobel GA (2002) Rainforest endophytes and bioactive products. Crit Rev Biotechnol 22: 315–333. doi: 10.1080/07388550290789531
|
| [23] |
Shoresh M, Yedidia I, Chet I (2005) Involvement of jasmonic acid/ethylenesignaling pathway in the systemic resistance induced in cucumber by Trichoderma asperellum T203. Phytopathology 95: 76–84. doi: 10.1094/PHYTO-95-0076
|
| [24] | Glick BR (2015) Beneficial Plant-Bacterial Interactions, Heidelberg: Springer, 243. |
| [25] |
Santoyo G, Moreno-Hagelsieb G, Del COM, et al. (2016) Plant growth-promoting bacterial endophytes. Microbiol Res 183: 92–99. doi: 10.1016/j.micres.2015.11.008
|
| [26] | Cerozi BDS, Fitzsimmons K (2016) Use of Bacillus spp. to enhance phosphorus vailability and serve as a plant growth promoter in aquaponics systems. Sci Hortic -Amsterdam 211: 277–282. |
| [27] |
Vinale F, Sivasithamparam K, Ghisalberti EL, et al. (2008) A novel role for Trichoderma secondary metabolites in the interactions with plants. Physiol Mol Plant P 72: 80–86. doi: 10.1016/j.pmpp.2008.05.005
|
| [28] |
Larsen J, Pineda-Sánchez H, Delgado-Arellano I, et al. (2017) Interactions between microbial plant growth promoters and their effects on maize growth performance in different mineral and organic fertilization scenarios. Rhizosphere 3: 75–81. doi: 10.1016/j.rhisph.2017.01.003
|
| [29] |
Kuklinsky-Sobral J, Araújo WL, Mendes R, et al. (2004) Isolation and characterization of soybean-associated bacteria and their potential for plant growth promotion. Environ Microbiol 6: 1244–1251. doi: 10.1111/j.1462-2920.2004.00658.x
|
| [30] |
Meng Q, Jiang H, Hao JJ (2016) Effects of Bacillus velezensis strain BAC03 in promoting plant growth. Biol Control 98: 18–26. doi: 10.1016/j.biocontrol.2016.03.010
|
| [31] |
Cohen AC, Travaglia CN, Bottini R, et al. (2009) Participation of abscisic acid and gibberellins produced by endophytic Azospirillum in the alleviation of drought effects in maize. Botany 87: 455–462. doi: 10.1139/B09-023
|
| [32] |
Silva HSA, Tozzi JPL, Terrasan CRF, et al. (2012) Endophytic microorganisms from coffee tissues as plant growth promoters and biocontrol agents of coffee leaf rust. Biol Control 63: 62–67. doi: 10.1016/j.biocontrol.2012.06.005
|
| [33] | Miransari M (2010) Current research, technology and education topics in applied microbiology and microbial biotechnology, In: Microbiology Book Series-2010 Edition, Spain. |
| [34] |
Vessey JK (2003) Plant growth promoting rhizobacteria as biofertilizers. Plant Soil 255: 571–586. doi: 10.1023/A:1026037216893
|
| [35] |
Richardson AE, Barea JM, Mcneill AM, et al. (2009) Acquisition of phosphorus and nitrogen in the rhizosphere and plant growth promotion by microorganisms. Plant Soil 321: 305–339. doi: 10.1007/s11104-009-9895-2
|
| [36] |
Rodriguez H, Fraga R (1999) Phosphate solubilizing bacteria and their role in plant growth promotion. Biotechnol Adv 17: 319–339. doi: 10.1016/S0734-9750(99)00014-2
|
| [37] |
Sturz AV, Nowak J (2000) Endophytic communities of rhizobacteria and the strategies required to create yield enhancing associations with crops. Appl Soil Ecol 15: 183–190. doi: 10.1016/S0929-1393(00)00094-9
|
| [38] |
Sudhakar P, Chattopadhyay GN, Gangwar SK, et al. (2000) Effect of foliar application of Azotobacter, Azospirillum and Beijerinckia on leaf yield and quality of mulberry (Morus alba). J Agric Sci 134: 227–234. doi: 10.1017/S0021859699007376
|
| [39] |
Karlidag H, Esitken A, Turan M, et al. (2007) Effects of root inoculation of plant growth promoting rhizobacteria (PGPR) on yield, growth and nutrient element contents of leaves of apple. Sci Hortic-Amsterdam 114: 16–20. doi: 10.1016/j.scienta.2007.04.013
|
| [40] | Ilyas N, Bano A (2012) Potencial use of soil microbial community in agriculture, In: Bacteria in Agrobiology: Plant Probiotics, 1 Eds., Berlin: Springer Berlin Heidelberg, 45–64. |
| [41] | Glick BR (2012) Plant growth-promoting bacteria: mechanisms and applications. Scientifica. |
| [42] |
Glick BR (2014) Bacteria with ACC deaminase can promote plant growth and help to feed the world. Microbiol Res 169: 30–39. doi: 10.1016/j.micres.2013.09.009
|
| [43] |
Brígido C, Glick BB, Oliveira S (2017) Survey of plant growth-promoting mechanisms in native Portuguese Chickpea Mesorhizobium isolates. Microbial Ecol 73: 900–915. doi: 10.1007/s00248-016-0891-9
|
| [44] |
Bhattacharyya PN, Jha DK (2012) Plant growth-promoting rhizobacteria (PGPR): emergence in agriculture. World J Microb Biotechnol 28: 1327–1350. doi: 10.1007/s11274-011-0979-9
|
| [45] |
Gray EJ, Smith DL (2005) Intracellular and extracellular PGPR: commonalities and distinctions in the plant-bacterium signaling processes. Soil Biol Biochem 37: 395–412. doi: 10.1016/j.soilbio.2004.08.030
|
| [46] | Figueiredo MVB, Martinez CR, Burity HA, et al. (2008) Plant growth-promoting rhizobacteria for improving nodulation and nitrogen fixation in the common bean (Phaseolus vulgaris L.). World J Microb Biotechnol 24: 1187e93. |
| [47] | Ahemad M, Kibret M (2014) Mechanisms and applications of plant growth promoting rhizobacteria: Current perspective. J King Saud Univ-Sci 26: 1–20. |
| [48] | Antoun H, Prévost D (2006) Ecology of Plant Growth Promoting Rhizobacteria, In: Siddiqui ZA, Editor, PGPR: Biocontrol and Biofertilization, Dordrecht: Springer Netherlands, 1–38. |
| [49] |
Brunner SM, Goos RJ, Swenson SJ, et al. (2015) Impact of nitrogen fixing and plant growth-promoting bacteria on a phloem-feeding soybean herbivore. Appl Soil Ecol 86: 71–81. doi: 10.1016/j.apsoil.2014.10.007
|
| [50] |
Smith SE, Manjarrez M, Stonor R, et al. (2015) Indigenous arbuscular mycorrhizal (AM) fungi contribute to wheat phosphate uptake in a semi-arid field environment, shown by tracking with radioactive phosphorus. Appl Soil Ecol 96: 68–74. doi: 10.1016/j.apsoil.2015.07.002
|
| [51] |
Mensah JA, Koch AM, Antunes PM, et al. (2015) High functional diversity within species of arbuscular mycorrhizal fungi is associated with differences in phosphate and nitrogen uptake and fungal phosphate metabolism. Mycorrhiza 25: 533–546. doi: 10.1007/s00572-015-0631-x
|
| [52] |
Augé RM, Toler HD, Saxton AM (2015) Arbuscular mycorrhizal symbiosis alters stomatal conductance of host plants more under drought than under amply watered conditions: a meta-analysis. Mycorrhiza 25: 13–24. doi: 10.1007/s00572-014-0585-4
|
| [53] |
Schübler A, Schwarzott D, Walker C (2001) A new fungal phylum, the Glomeromycota: phylogeny and evolution. Mycol Res 105: 1413–1421. doi: 10.1017/S0953756201005196
|
| [54] |
Dodd JC, Boddington CL, Rodriguez A, et al. (2000) Mycelium of Arbuscular Mycorrhizal fungi (AMF) from different genera: form, function and detection. Plant Soil 226: 131–151. doi: 10.1023/A:1026574828169
|
| [55] |
Artursson V, Finlay RD, Jansson JK (2006) Interactions between arbuscular mycorrhizal fungi and bacteria and their potential for stimulating plant growth. Environ Microbiol 8: 1–10. doi: 10.1111/j.1462-2920.2005.00942.x
|
| [56] |
Miransari M (2011) Interactions between arbuscular mycorrhizal fungi and soil bacteria. Appl Microbiol Biotechnol 89: 917–930. doi: 10.1007/s00253-010-3004-6
|
| [57] |
Zhang L, Xu M, Liu Y, et al. (2016) Carbon and phosphorus exchange may enable cooperation between an arbuscular mycorrhizal fungus and a phosphate-solubilizing bacterium. New Phytologist 210: 1022–1032. doi: 10.1111/nph.13838
|
| [58] |
Chaiyasen A, Young JPW, Teaumroong N, et al. (2014) Characterization of arbuscular mycorrhizal fungus communities of Aquilaria crassna and Tectona grandis roots and soils in Thailand plantations. Plos One 9: e112591. doi: 10.1371/journal.pone.0112591
|
| [59] | Chaiyasen A, Douds DD, Gavinlertvatan P, et al. (2017) Diversity of arbuscular mycorrhizal fungi in Tectona grandis Linn.f. plantations and their effects on growth of micropropagated plantlets. New Forest 48: 547–562. |
| [60] | Brundrett MC, Bougher N, Dell B, et al. (1996) Working with mycorrhizas in forestry and agriculture. |
| [61] | Smith S, Read D (2008) Mycorrhizal Symbiosis, New York: Academic Press. |
| [62] |
Porcel R, Ruiz-Lozano JM (2004) Arbuscular mycorrhizal influence on leaf water potential, solute accumulation, and oxidative stress in soybean plants subjected to drought stress. J Exp Bot 55: 1743–1750. doi: 10.1093/jxb/erh188
|
| [63] |
Doubková P, Vlasáková E, Sudová R (2013) Arbuscular mycorrhizal symbiosis alleviates drought stress imposed on Knautia arvensis plants in serpentine soil. Plant Soil 370: 149–161. doi: 10.1007/s11104-013-1610-7
|
| [64] | Azcón-Aguilar C, Barea JM (1996) Arbuscular mycorrhizas and biological control of soil-borne plant pathogens-an overview of the mechanisms involved. Mycorrhiza 6: 457–464. |
| [65] |
Wright SF, Upadhyaya A (1998) A survey of soils for aggregate stability and glomalin, a glycoprotein produced by hyphae of arbuscular mycorrhizal fungi. Plant Soil 198: 97–107. doi: 10.1023/A:1004347701584
|
| [66] |
Rillig MC (2004) Arbuscular mycorrhizae, glomalin, and soil aggregation. Can J Soil Sci 84: 355–363. doi: 10.4141/S04-003
|
| [67] |
Rillig MC, Ramsey PW, Morris S, et al. (2003) Glomalin, an arbuscularmycorrhizal fungal soil protein, responds to land-use change. Plant Soil 253: 293–299. doi: 10.1023/A:1024807820579
|
| [68] |
Bhardwaj D, Ansari MW, Sahoo K, et al. (2014) Biofertilizers function as key player in sustainable agriculture by improving soil fertility, plant tolerance and crop productivity. Microb Cell Fact 13: 1–10. doi: 10.1186/1475-2859-13-1
|
| [69] | Goswami D, Thakker JN, Dhandhukia PC (2016) Portraying mechanics of plant growth promoting rhizobacteria (PGPR): A review. Cogent Food Agr 2: 1–19. |
| [70] |
Kang SM, Khan AL, Waqas M, et al. (2014) Plant growth-promoting rhizobacteria reduce adverse effects of salinity and osmotic stress by regulating phytohormones and antioxidants in Cucumis sativus. J Plant Interact 9: 673–682. doi: 10.1080/17429145.2014.894587
|
| [71] |
Bartels D, Sunkar R (2005) Drought and salt tolerance in plants. Crit Rev Plant Sci 24: 23–58. doi: 10.1080/07352680590910410
|
| [72] | Hussain TM, Chandrasekhar T, Hazara M, et al. (2008) Recent advances in salt stress biology-A review. Biotechnol Mol Biol Rev 3: 8–13. |
| [73] |
Saleem M, Arshad M, Hussain S, et al. (2007) Perspective of plant growth promoting rhizobacteria (PGPR) containing ACC deaminase in stress agriculture. J Ind Microbiol Biotechnol 34: 635–648. doi: 10.1007/s10295-007-0240-6
|
| [74] | Shaharoona B, Arshad M, Zahir ZA (2006) Performance of Pseudomonas spp. containing ACC-deaminase for improving growth and yield of maize (Zea mays L.) in the presence of nitrogenous fertilizer. Soil Biol Biochem 38: 2971–2975. |
| [75] |
Egamberdiyeva D (2007) The effect of plant growth promoting bacteria on growth and nutrient uptake of maize in two different soils. Appl Soil Ecol 36: 184–189. doi: 10.1016/j.apsoil.2007.02.005
|
| [76] | Barriuso J, Solano BR, Gutiérrez MFJ (2008) Protection against pathogen and salt stress by four plant growth-promoting Rhizobacteria isolated from Pinus sp. on Arabidopsis thaliana. Biol Control 98: 666–672. |
| [77] |
Kohler J, Hernández JA, Caravaca F, et al. (2009) Induction of antioxidant enzymes is involved in the greater effectiveness of a PGPR versus AM fungi with respect to increasing the tolerance of lettuce to severe salt stress. Environ Exp Botany 65: 245–252. doi: 10.1016/j.envexpbot.2008.09.008
|
| [78] | Sandhya V, Ali SZ, Grover M, et al. (2010) Effect of plant growth promoting Pseudomonas spp. on compatible solutes, antioxidant status and plant growth of maize under drought stress. Plant Growth Regul 62: 21–30. |
| [79] |
Seneviratne G, Jayasekara APDA, De Silva MSDL, et al. (2011) Developed microbial biofilms can restore deteriorated convencional agricultural soils. Soil Biol Biochem 43: 1059–1062. doi: 10.1016/j.soilbio.2011.01.026
|
| [80] | Shukla PS, Agarwal PK, Jha B (2012) Improved salinity tolerance of Arachis hypogaea (L.) by the interaction of halotolerant plant-growth-promoting-rhizobacteria. J Plant Growth Regul 31: 195–206. |
| [81] |
Swarnalakshmi K, Prasanna R, Kumar A, et al. (2013) Evaluating the influence of novel cyanobacterial biofilmed biofertilizers on soil fertility and plant nutrition in wheat. Eur J Soil Biol 55: 107–116. doi: 10.1016/j.ejsobi.2012.12.008
|
| [82] |
Sarma RK, Saikia R (2014) Alleviation of drought stress in mug bean by strain Pseudomonas aeruginosa GGRJ21. Plant Soil 377: 111–126. doi: 10.1007/s11104-013-1981-9
|
| [83] | Cardinale M, Ratering S, Suarez C, et al. (2015) Paradox of plant growth promotion potencial of rhizobacteria and their actual promotion effect on growth of barley (Hordeum vulgare L.) under salt stress. Microbiol Res 181: 22–32. |
| [84] |
Islam F, Yasmeen T, Arif MS, et al. (2016) Plant growth promotion bacteria confer salt tolerance in Vigna radiata by up-regulating antioxidant defense and biological soil fertility. Plant Growth Regul 80: 23–36. doi: 10.1007/s10725-015-0142-y
|
| [85] |
Shahzad R, Khan AL, Bilal S, et al. (2017) Inoculation of abscisic acid-producing endophytic bacteria enhances salinity stress tolerance in Oryza sativa. Environ Exp Botany 136: 68–77. doi: 10.1016/j.envexpbot.2017.01.010
|
| [86] |
Rajkumar M, Sandhya S, Prasad MNV, et al. (2012) Perspectives of plant-associated microbes in heavy metal phytoremediation. Biotechnol Adv 30: 1562–1574. doi: 10.1016/j.biotechadv.2012.04.011
|
| [87] | Saleh S, Huang XD, Greenberg BM, et al. (2004) Phytoremediation of persistent organic contaminants in the environment, In: Singh A, Ward O, Editors, Appl ied Bioremediation Phytoremediation, Berlin: Springer-Verlag, 115–134. |
| [88] |
Zhuang X, Chen J, Shim H, et al. (2007) New advances in plant growth-promotion rhizobacteria for bioremediation. Environ Int 33: 406–413. doi: 10.1016/j.envint.2006.12.005
|
| [89] | Ma Y, Prasad MNV, Rajkumar M, et al. (2010) Plant growth promoting rhizobacteria and endophytes accelerate phytoremediation of metalliferous soils. Biotechnol Adv 29: 248–258. |
| [90] |
Pinter IF, Salomon MV, Berli F, et al. (2017) Characterization of the As(III) tolerance conferred by plant growth promoting rhizobacteria to in vitro-grown grapevine. Appl Soil Ecol 109: 60–68. doi: 10.1016/j.apsoil.2016.10.003
|
| [91] | Babu AG, Kim JD, Oh BT (2013) Enhancement of heavy metal phytoremediation by Alnus firma with endophytic Bacillus thuringiensis GDB-1. J Hazard Mat 250: 477–483. |
| [92] | Kuklinsky-Sobral J, Araújo WL, Mendes R, et al. (2004) Isolation and characterization of soybean-associated bacteria and their potential for plant growth promotion. Environ Microbiol 12: 1244–1251. |
| [93] |
Chelius MK, Triplett EW (2001) The diversity of archaea and bacteria in association with the roots of Zea mays L. Microb Ecol 41: 252–263. doi: 10.1007/s002480000087
|
| [94] |
Garbeva P, van Overbeek LS, van Vuurde JEL, et al. (2001) Analysis of endophytic bacterial communities of potato by plating and denaturing gradient gel electrophoresis (DGGE) of 16S rDNA based PCR fragments. Microb Ecol 41: 369–383. doi: 10.1007/s002480000096
|
| [95] |
Araújo WL, Marcon J, Maccheroni WJ, et al. (2002) Diversity of endophytic bacterial populations and their interaction with Xylella fastidiosa in citrus plants. Appl Environ Microbiol 68: 4906–4914. doi: 10.1128/AEM.68.10.4906-4914.2002
|
| [96] |
Zinniel DK, Lambrecht P, Harris B, et al. (2002) Isolation and characterization of endophytic colonizing bacteria from agronomic crops and prairie plants. Appl Environ Microbiol 68: 2198–2208. doi: 10.1128/AEM.68.5.2198-2208.2002
|
| [97] |
Idris R, Trifonova R, Puschenreiter M, et al. (2004) Bacterial communities associated with flowering plants of the Ni Hyperaccumulator Thlaspi goesingense. Appl Environ Microbiol 70: 2667–2677. doi: 10.1128/AEM.70.5.2667-2677.2004
|
| [98] |
Andreote FD, Carneiro RT, Salles JF, et al. (2009) Culture-independent assessment of Alphaproteobacteria related to order Rhizobiales and the diversity of cultivated Methylobacterium in the rhizosphere and rhizoplane of transgenic eucalyptus. Microb Ecol 57: 82–93. doi: 10.1007/s00248-008-9405-8
|
| [99] |
Sagaram US, DeAngelis KM, Trivedi P, et al. (2009) Bacterial diversity analysis of Huanglongbing pathogen-infected citrus, using phylochip arrays and 16S rRNA gene clone library sequencing. Appl Environ Microbiol 75: 1566–1574. doi: 10.1128/AEM.02404-08
|
| [100] |
Gv DMP, Magalhães KT, Lorenzetii ER, et al. (2012) A multiphasic approach for the identification of endophytic bacterial in strawberry fruit and their potential for plant growth promotion. Microb Ecol 63: 405–417. doi: 10.1007/s00248-011-9919-3
|
| [101] |
Xia X, Lie TK, Qian X (2011) Species diversity, distribution, and genetic structure of endophytic and epiphytic Trichoderma associated with banana roots. Microb Ecol 61: 619–625. doi: 10.1007/s00248-010-9770-y
|
| [102] |
Ikeda AC, Bassani LL, Adamoski D, et al. (2013) Morphological and genetic characterization of endophytic bacteria isolated from roots of different maize genotypes. Microb Ecol 65: 154–160. doi: 10.1007/s00248-012-0104-0
|
| [103] | Verma VC, Gond SK, Kumar A, et al. (2009) Endophytic actinomycetes from Azadirachta indica A. Juss.: isolation, diversity, and anti-microbial activity. Microb Ecol 57: 749–756. |
| [104] |
Manter DK, Delgado JA, Holm DG, et al. (2010) Pyrosequencing reveals a highly diverse and cultivar-specific bacterial endophyte community in potato roots. Microb Ecol 60: 157–166. doi: 10.1007/s00248-010-9658-x
|
| [105] | Cruz LM, Souza EM, Weber OB, et al. (2001) 16S Ribosomal DNA characterization of nitrogen-fixing bacteria isolated from banana (Musa spp.) and pineapple (Ananas comosus (L.) Merril). Appl Environ Microbiol 67: 2375–2379. |
| [106] |
Marques ASA, Marchaison A, Gardan L, et al. (2008) BOXPCR-based identification of bacterial species belonging to Pseudomonas syringae: P. viridiflava group. Gen Mol Biol 31: 106–115. doi: 10.1590/S1415-47572008000100019
|
| [107] |
Menna P, Barcellos FG, Hungria M (2009) Phylogeny and taxonomy of a diverse collection of Bradyrhizobium strains based on multilocus sequence analysis of 16S rRNA, ITS, glnII, recA, atpD and dnaK genes. Int J Syst Evol Microbiol 59: 2934–2950. doi: 10.1099/ijs.0.009779-0
|
| [108] |
Zhang XX, Gao JS, Cao YH (2013) Long-term rice and green manure rotation alters the endophytic bacterial communities of the rice root. Microb Ecol 66: 917–926. doi: 10.1007/s00248-013-0293-1
|
| [109] |
Onstott TC, Phelps TJ, Colwell FS, et al. (1998) Observations pertaining to the origin and ecology of microorganisms recovered from the deep subsurface of Taylorsville Basin, Virginia. Geomicrobiol J 15: 353–385. doi: 10.1080/01490459809378088
|
| [110] |
Nadkarni MA, Martin FE, Hunter N, et al. (2009) Methods for optimizing DNA extraction before quantifying oral bacterial numbers by real-time PCR. FEMS Microbiol Lett 296: 45–51. doi: 10.1111/j.1574-6968.2009.01629.x
|
| [111] | Muyzer G, De WEC, Uitterlinden AG (1993) Profiling of complex microbial populations by denaturing gradient gel electroforesis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Appl Environ Microbiol 59: 695–700. |
| [112] |
Orphan VJ, Taylor LT, Hafenbradl D, et al. (2000) Culture-dependent and culture-independent characterization of microbial assemblages associated with high-temperature petroleum reservoirs. Appl Environ Microbiol 66: 700–711. doi: 10.1128/AEM.66.2.700-711.2000
|
| [113] | Monteiro JM, Vollú RE, Coelho MRR, et al. (2009) Comparison of the bacterial community and characterization of plant growth-promoting rhizobacteria from different genotypes of Chrysopogon zizanioides (L.) Roberty (Vetiver) rhizospheres. J Microbiol 4: 363–370. |
| [114] | Sun L, Qiu F, Zhang X, et al. (2008) Endophytic bacterial diversity in rice (Oryza sativa L.) roots estimated by 16S rDNA sequence analysis. Microb Ecol 55: 415–424. |
| [115] | Snyder LAS, Loman N, Pallen MJ, et al. (2009) Next-generation sequencing-the promise and perils of charting the great microbial unknown. Microb Ecol 57: 1–3. |
| [116] |
Studholme DJ, Glover RH, Boonham N (2011) Application of high-throughput DNA sequencing in phytopathology. Annu Rev Phytopathol 49: 87–105. doi: 10.1146/annurev-phyto-072910-095408
|
| [117] |
Lundberg DS, Lebeis SL, Paredes SH (2012) Defining the core Arabidopsis thaliana root microbiome. Nature 488: 86–90. doi: 10.1038/nature11237
|
| [118] | Akinsanya MA, Goh JK, Lim SP, et al. (2015) Metagenomics study of endophytic bacteria in Aloe vera using next-generation technology: Genom Data 6: 159–163. |
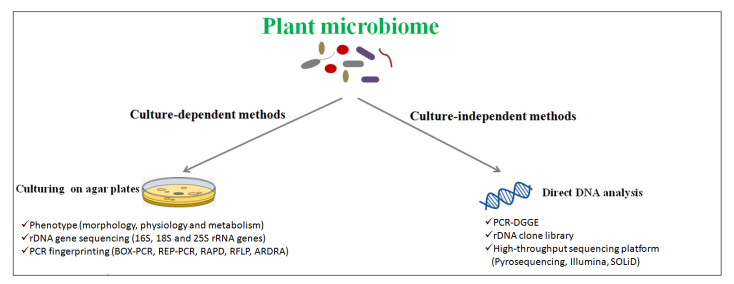
Figures(1) / Tables(3)

Luciana Porto de Souza Vandenberghe, Lina Marcela Blandon Garcia, Cristine Rodrigues, Marcela Cândido Camara, Gilberto Vinícius de Melo Pereira, Juliana de Oliveira, Carlos Ricardo Soccol. Potential applications of plant probiotic microorganisms in agriculture and forestry[J]. AIMS Microbiology, 2017, 3(3): 629-648. doi: 10.3934/microbiol.2017.3.629







 DownLoad:
DownLoad: