Citation: Karina Sałek, Tony Gutierrez. Surface-active biopolymers from marine bacteria for potential biotechnological applications[J]. AIMS Microbiology, 2016, 2(2): 92-107. doi: 10.3934/microbiol.2016.2.92

| [1] |
Banat IM, Makkar RS, Cameotra SS (2000) Potential commercial applications of microbial surfactants. Appl Microbiol Biotechnol 53: 495–508. doi: 10.1007/s002530051648

|
| [2] | Desai JD, Banat IM (1997) Microbial production of surfactants and their commercial potential. Microbiol Molec Biol Rev 61: 47–64. |
| [3] |
Singh P, Cameotra SS (2004) Potential applications of microbial surfactants in biomedical sciences. Trends Biotechnol 22: 142–146. doi: 10.1016/j.tibtech.2004.01.010
|
| [4] |
Banat IM, Franzetti A, Gandolfi I, et al. (2010) Microbial biosurfactants production, applications and future potential. Appl Microbiol Biotechnol 87: 427–444. doi: 10.1007/s00253-010-2589-0
|
| [5] |
Campos JM, Stamford TL, Sarubbo LA, et al. (2013) Microbial biosurfactants as additives for food industries: a review. Biotechnol Progr 29: 1097–1108. doi: 10.1002/btpr.1796
|
| [6] | Fracchia L, Cavallo M, Martinotti M, et al. (2012) Biosurfactants and bioemulsifiers biomedical and related applications – present status and future potentials. Biomedical Science, Engineering and Technology, Chapter 14, 325–370. |
| [7] |
Lourith N, Kanlayavattanakul M (2009) Natural surfactants used in cosmetics: glycolipids. Int J Cosmetic Sci 31:255–261. doi: 10.1111/j.1468-2494.2009.00493.x
|
| [8] | Senkhon KK, Khanna S, Cameotra SS (2012) Biosurfactant production and potential correlation with esterase activity. J Pet Environ Biotechnol 3: 133. |
| [9] |
Dreja M, Vockenroth I, Plath N (2012) Biosurfactants – exotic specialties or ready for application? Tenside Surf Det 49: 10–17. doi: 10.3139/113.110158
|
| [10] | Poremba K, Gunkel W, Lang S, et al. (1991) Marine biosurfactants, III. Toxicity testing with marine microorganisms and comparison with synthetic surfactants. Z Naturforsch C 46: 210–216. |
| [11] | Marchant R, Banat IM (2012a) Microbial biosurfactants: challenges and opportunities for future exploitation. Trends Biotechnol 30: 558–565. |
| [12] | Marchant R, Banat IM (2012b) Biosurfactants: a sustainable replacement for chemical surfactants? Biotechnol Lett 34: 1597–1605. |
| [13] |
Satpute SK, Banat IM, Dhakephalkar PK, et al. (2010) Biosurfactants, bioemulsifiers and exopolysaccharides from marine microorganisms. Biotechnol Adv 28: 436–450. doi: 10.1016/j.biotechadv.2010.02.006
|
| [14] |
Shekhar S, Sundaramanickam A, Balasubramanian T (2015) Biosurfactant producing microbes and their potential applications: a review. Critical Rev Environ Sci Technol 45: 1522–1554. doi: 10.1080/10643389.2014.955631
|
| [15] | Kronberg B, Holmberg K, Lindman B (2014) Surface chemistry of surfactants and polymers. John Wiley & Sons, Ltd. |
| [16] |
Rosenberg E, Ron EZ (1997) Bioemulsans: microbial polymeric emulsifiers. Curr Opin Biotechnol 8: 313–316. doi: 10.1016/S0958-1669(97)80009-2
|
| [17] | Klekner V, Kosaric N (1993) Biosurfactants for cosmetic. Surfactant Sci Ser 48: 373–389. |
| [18] |
Shepherd R, Rockey J, Sutherland IW, et al. (1995) Novel bioemulsifiers from microorganisms for use in foods. J Biotechnol 40: 207–217. doi: 10.1016/0168-1656(95)00053-S
|
| [19] |
Garti N (1999) What can nature offer from an emulsifier point of view: trends and progress? Colloids Surf 152: 125–146. doi: 10.1016/S0927-7757(98)00621-9
|
| [20] | Hasenhuettle GL, Hartel RW (1997) Food emulsifiers and their applications. Chapman and Hall, New York. |
| [21] |
Hosono A, Lee J, Ametani A, et al. (1997) Characterization of a water-soluble polysaccharide fraction with immunopotentiating activity from Bifidobacterium adolescentris M101-4. Biosci Biotechnol Biochem 61: 312–316. doi: 10.1271/bbb.61.312
|
| [22] |
Nakajima H, Suzuki Y, Kaizu H, et al (1992) Cholesterol-lowering activity of ropy fermented milk. J Food Sci 57: 1327–1329. doi: 10.1111/j.1365-2621.1992.tb06848.x
|
| [23] |
Ruijssenaars HJ, Stingele F, Hartmans S (2000) Biodegradability of food-associated extracellular polysaccharides. Curr Microbiol 40: 194–199. doi: 10.1007/s002849910039
|
| [24] | Sanderson GR (1990) The functional properties and application of microbial polysaccharides – a supplier’s view. In: Phillips GO, Wedlock DJ, Williams PA, Editor, Gums and stabilizers for the food industry, Oxford: IRL Press, 5: 333–339. |
| [25] | Dickinson E, Murray BS, Stainsby G (1988) Protein adsorption at air-water and oil-water interfaces. In: Dickinson E, Stainsby G, Editor, Advances in food emulsions and foams. London: Elsevier, 123–162. |
| [26] |
Randall RC, Phillips GO, Williams PA (1988) The role of the proteinaceous component on the emulsifying properties of gum arabic. Food Hydrocoll 2: 131–140. doi: 10.1016/S0268-005X(88)80011-0
|
| [27] | Sałek K, Zgoła-Grześkowiak A, Kaczorek E (2013) Modification of surface and enzymatic properties of Achromobacter denitrificans and Stenotrophomonas maltophilia in association with diesel oil biodegradation enhanced with alkyl polyglucosides. Colloids Surf B 111: 36–42. |
| [28] |
Ryan LD, Kaler EW (2001) Alkyl polyglucoside microemulsion phase behavior. Colloids Surf A Physicochem Eng Asp 176: 69–83. doi: 10.1016/S0927-7757(00)00614-2
|
| [29] |
Holmberg K (2001) Natural surfactants. Curr Opin Colloid In 6: 148–159. doi: 10.1016/S1359-0294(01)00074-7
|
| [30] | The European Parliament and The Council Of The European Union (2004) Regulation (EC) No 648/2004 of the European Parliament and of the Council of 31 March 2004 on detergents, Official Journal of the European Union. |
| [31] |
Hansell DA, Carlson CA (1998) Deep-ocean gradients in the concentration of dissolved organic carbon. Nature 395: 263–268. doi: 10.1038/26200
|
| [32] |
Chin W-C, Orellana MV, Verdugo P (1998) Formation of microgels by spontaneous assembly of dissolved marine polymers. Nature 391: 568–572. doi: 10.1038/35345
|
| [33] | Verdugo P (1994) Polymer gel phase transition in condensation-decondensation of secretory products. Adv Polymer Sci 110: 145–156. |
| [34] |
Azam F (1998) Microbial control of oceanic carbon flux: the plot thickens. Science 280: 694–696. doi: 10.1126/science.280.5364.694
|
| [35] | Decho AW (1990) Microbial exopolymer secretions in ocean environments: their role(s) in food webs and marine processes. In: Barnes M, Editor, Oceanography marine biology annual review. Aberdeen: Aberdeen University Press, 73–153. |
| [36] | Santschi PH, Guo L, Means JC, et al. (1998) Natural organic matter binding of trace metal and trace organic contaminants in estuaries. In: Bianchi TS, Pennock JR, Twilley R, Editor, Biogeochemistry of Gulf of Mexico Estuaries. New York: Wiley, 347–380. |
| [37] | Bhaskar PV, Bhosle NB (2005) Microbial extracellular polymeric substances in marine biogeochemical processes. Curr Sci 88: 45–53. |
| [38] | Ford T, Sacco E, Black J, et al. (1991) Characterization of exopolymers of aquatic bacteria by pyrolysis-mass spectrometry. Appl Environ Microbiol 57: 1595–1601. |
| [39] | Kennedy AFD, Sutherland IW (1987) Analysis of bacterial exopolysaccharides. Biotechnol Appl Biochem 9: 12–19. |
| [40] | Thavasi R, Banat IM (2014) Biosurfactant and bioemulsifiers from marine sources. Mulligan CN, Sharma SK, Mudhoo A, Editor, Hardback: CRC Press, chapter 5, 125–146. |
| [41] |
Gutierrez T, Berry D, Yang T, et al. (2013) Role of bacterial exopolysaccharides (EPS) in the fate of the oil released during the Deepwater Horizon oil spill. PLoS ONE, 8: e67717. doi: 10.1371/journal.pone.0067717
|
| [42] | Gutierrez T, Biller D, Shimmield T, et al. (2012) Metal binding properties of the EPS produced by Halomonas sp. TG39 and its potential in enhancing trace element bioavailability to eukaryotic phytoplankton. BioMetals 25: 1185–1194. |
| [43] |
Gutierrez T, Shimmield T, Haidon C, et al. (2008) Emulsifying and metal ion binding activity of a glycoprotein exopolymer produced by Pseudoalteromonas species TG12. Appl Environ Microbiol 74: 4867–4876. doi: 10.1128/AEM.00316-08
|
| [44] | Thavasi R, Jayalakshmi S, Banat IM (2011) Biosurfactant from marine bacterial isolates. In Current Research Technology and Education Topics in Applied Microbiology and Microbial Biotechnology Book Series. Mendez-Vilas A, Editor, Badajoz: Formatex Research Center, 2: 1367–1373. |
| [45] | Gutierrez T, Morris G, Green DH (2009) Yield and physicochemical properties of EPS from Halomonas sp. strain TG39 identifies a role for protein and anionic residues (sulphate and phosphate) in emulsification of n-hexadecane. Biotechnol Bioeng 103: 207–216. |
| [46] |
Belsky I, Gutnick DL, Rosenberg E (1979) Emulsifier of Arthrobacter RAG-1: determination of emulsifier-bound fatty acids. FEBS Lett 101: 175–178. doi: 10.1016/0014-5793(79)81320-4
|
| [47] | Garti N, Leser ME (1999) Natural hydrocolloids as food emulsifiers. In: Karsa DR, Editor, Design and selection of performance surfactants. Sheffield, UK: Sheffield Academic Press, 104–145. |
| [48] | Kaplan N, Zosim Z, Rosenberg E (1987) Reconstitution of emulsifying activity of Acinetobacter calcoaceticus BD4 emulsan by using pure polysaccharide and protein. Appl Environ Microbiol 53: 440–446. |
| [49] | Franzetti A, Gandolfi I, Fracchia L, et al. (2014) Biosurfactant use in heavy metal removal from industrial effluents and contaminated sites. In: Biosurfactants: Production and Utilization – Processes, Technologies and Economics. Kosaric N, Sukan FV, Editor, CRC Press, 361–369. |
| [50] | Franzetti A, Caredda P, Ruggeri C, et al. (2009) Potential applications of surface active compounds by Gordonia sp. Strain BS29 in soil remediation technologies. Chemosphere 75: 801–807. |
| [51] |
Asci Y, Nurbas M, Acikel YS (2010) Investigation of sorption/desorption equilibria of heavy metals ions on/from quartz using rhamnolipid biosurfactant. J Environ Manage 91: 724–731. doi: 10.1016/j.jenvman.2009.09.036
|
| [52] |
Verdugo P, Alldredge AL, Azam F, et al. (2004) The oceanic gel phase: a bridge in the DOM-POM continuum. Mar Chem 92: 67–85. doi: 10.1016/j.marchem.2004.06.017
|
| [53] |
Zosim Z, Gutnick D, Rosenberg E (1983) Uranium binding by emulsan and emulsanosols. Biotechnol Bioeng 25: 1725–1735. doi: 10.1002/bit.260250704
|
| [54] |
Ron E, Ronserberg E (2001) Natural roles of biosurfactants. Environ Microbiol 3:229–236. doi: 10.1046/j.1462-2920.2001.00190.x
|
| [55] |
Long RA, Azam F (1996) Abundant protein-containing particles in the sea. Aquat Microb Ecol 10: 213–221. doi: 10.3354/ame010213
|
| [56] |
Kato A (2002) Industrial applications of Maillard-type protein-polysaccharide conjugates. Food Sci Technol Res 8: 193–199. doi: 10.3136/fstr.8.193
|
| [57] |
Messner P (1997) Bacterial glycoproteins. Glycoconjugate J 14: 3–11. doi: 10.1023/A:1018551228663
|
| [58] |
Guezennec J (2002) Deep-sea hydrothermal vents: a new source of innovative bacterial exopolysaccharides of biotechnological interest? J Ind Microbiol Biotechnol 29: 204–208. doi: 10.1038/sj.jim.7000298
|
| [59] | Head IM, Jones MD, Roling WFM (2006) Marine microorganisms make a meal of oil. Nature 4: 173–182. |
| [60] | Andelman JB, Suess MJ (1970) Polynuclear aromatic hydrocarbons in the water environment. Bull World Health Organ 43: 479–508. |
| [61] | Gunnison D, Alexander M (1975) Basis for the resistance of several algae to microbial decomposition. Appl Microbiol 29: 729–738. |
| [62] | Gol’man LP, Mikhaseva MF, Reznikov VM. (1973). Infrared spectra of lignin preparations of pteridophytes and seaweeds. Dokl Akad Nauk BSSR 17: 1031–1033. |
| [63] |
Pastuska G (1961) Die Kieselgelschicht-Chromatographie von Phenolen und Phenolcarbensiuren. I Z Anal Chem 179: 355–358. doi: 10.1007/BF00462690
|
| [64] | Zelibor JL, Romankiw L, Hatcher PG, et al. (1988) Comparative analysis of the chemical composition of mixed and pure cultures of green algae and their decomposed residues by 13C nuclear magnetic resonance spectroscopy. Appl Environ Microbiol 54: 1051–1060. |
| [65] |
Binark N, Guven KC, Gezgin T, et al. (2000) Oil pollution of marine algae. Bull Environ Contamin Toxicol 64: 866–872. doi: 10.1007/s0012800083
|
| [66] |
Kowalewska G (1999) Phytoplankton – the main factor responsible for transport of polynuclear aromatic hydrocarbons from water to sediments in the Southern Baltic ecosystem. ICES J Mar Sci 56: 219–222. doi: 10.1006/jmsc.1999.0607
|
| [67] |
Buchan A, Collier LS, Neidle EL, et al. (2000) Key aromatic-ring-cleaving enzyme, protocatechuate 3,4-dioxygenase, in the ecologically important marine Roseobacter lineage. Appl Environ Microbiol 66: 4662– 4672. doi: 10.1128/AEM.66.11.4662-4672.2000
|
| [68] |
Buchan A, Neidle EL, Moran MA (2004) Diverse organization of genes of the β-ketoadipate pathway in members of the marine Roseobacter lineage. Appl Environ Microbiol 70: 1658–1668. doi: 10.1128/AEM.70.3.1658-1668.2004
|
| [69] |
Moran MA, Belas R, Schell MA, et al. (2007) Ecological genomics of marine Roseobacters. Appl Environ Microbiol 73: 4559–4569. doi: 10.1128/AEM.02580-06
|
| [70] |
Makkar R, Cameotra S, Banat IM (2011) Advances in utilization of renewable substrates for biosurfactant production. AMB Express, 1: 1–5. doi: 10.1186/2191-0855-1-1
|
| [71] |
Hauthal HG (2012) Biosurfactants, New Ingredients and Formulations, Sustainability, Forum for Innovations. Tenside Surf Det 49: 61–74. doi: 10.3139/113.110168
|
| [72] | Mukherjee S, Das P, Sen R (2006) Towards commercial production of microbial surfactants. Trends Biotechnol 24: 509–515. |
| [73] |
Develter DWG, Lauryssen LML (2010) Properties and industrial applications of sophorolipids. Eur J Lipid Sci Tech 112: 628–638. doi: 10.1002/ejlt.200900153
|
| [74] |
Arias S, del Moral A, Ferrer MR, et al. (2003) Mauran, an exopolysaccharide produced by the halophilic bacterium Halomonas maura, with a novel composition and interesting properties for biotechnology. Extremophiles 7: 319–326. doi: 10.1007/s00792-003-0325-8
|
| [75] | Garti N, Leser ME (1999) Natural hydrocolloids as food emulsifiers. In: Karsa DR, Editor. Design and selection of performance surfactants. Sheffield, UK: Sheffield Academic Press, 104–145. |
| [76] | Develter DWG, Fleurackers SJJ (2010) Surfactants from renewable resources. West Sussex: John Wiley & Sons, 213–238. |
| [77] |
Campos JM, Montenegro Stamford TL, Sarubbo LA, et al. (2013) Microbial biosurfactants as additives for food industries: a review. Biotechnol Progress 29: 1097–1108. doi: 10.1002/btpr.1796
|
| [78] |
Xu J, Shpak E, Gu T, et al. (2005) Production of recombinant plant gum with tobacco cell culture in bioreactor and gum characterization. Biotechnol Bioeng 90: 578–588. doi: 10.1002/bit.20441
|
| [79] |
Lawrence A, Balakrishnan M, Joseph TC, et al. (2014) Functional and molecular characterization of a lipopeptide surfactant from the marine sponge-associated eubacteria Bacillus licheniformis NIOT-AMKV06 of Andaman and Nicobar Islands, India. Mar Pollut Bull 82: 76–85. doi: 10.1016/j.marpolbul.2014.03.018
|
| [80] |
Hamman JH (2010) Chitosan based polyelectrolyte complexes as potential carrier materials in drug delivery systems. Mar Drugs 8: 1305–1322. doi: 10.3390/md8041305
|
| [81] |
Castro RG, Panilaitis B, Kaplan DL (2008) Emulsan, a tailorable biopolymer for controlled release. Biores Technol 99: 4566–4571. doi: 10.1016/j.biortech.2007.06.059
|
| [82] |
Abdel-Mawgoud AM, Lépine F, Déziel E (2010) Rhamnolipids: diversity of structures, microbial origins and roles. Appl Microbiol Biotechnol 86: 1323–1336. doi: 10.1007/s00253-010-2498-2
|
| [83] | Huang TT (2014) Carbohydrate esters as inducers for gene expression. Patent WO 2013003291 A2. |
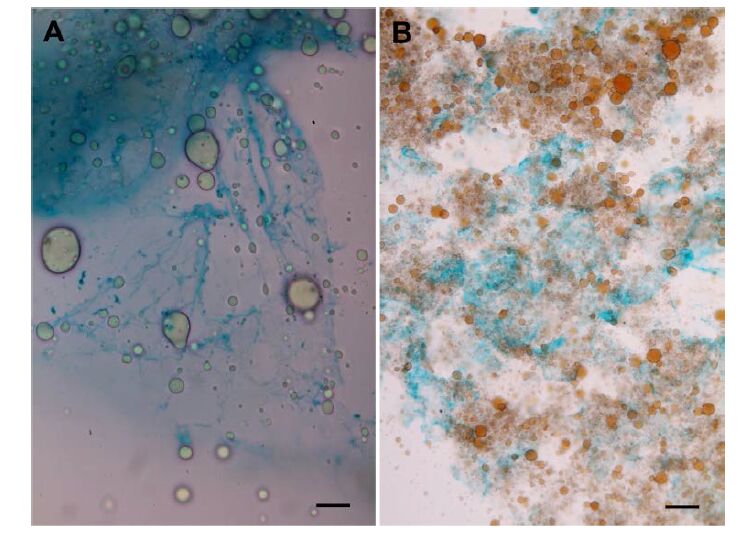
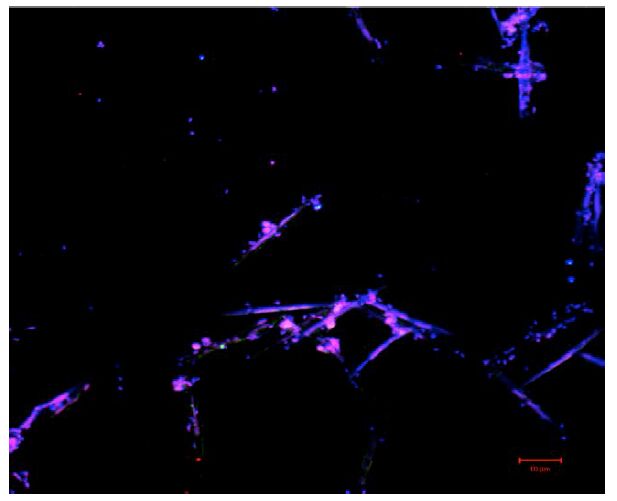
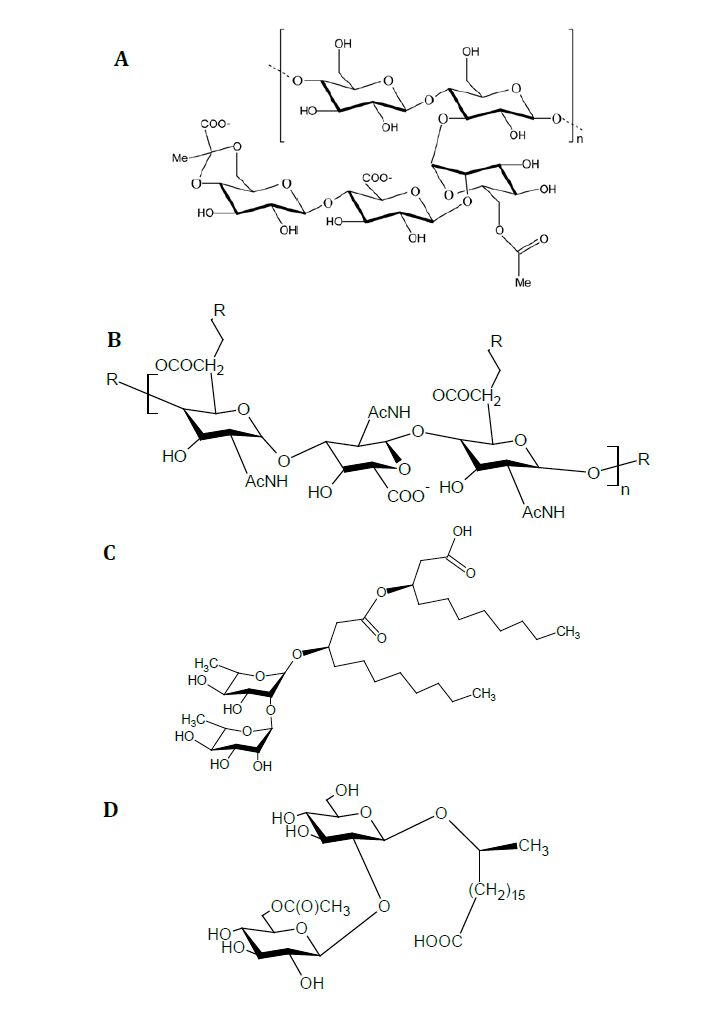
Figures(3)

Karina Sałek, Tony Gutierrez. Surface-active biopolymers from marine bacteria for potential biotechnological applications[J]. AIMS Microbiology, 2016, 2(2): 92-107. doi: 10.3934/microbiol.2016.2.92







 DownLoad:
DownLoad: