Multiple sclerosis (MS), is a debilitating neurological disease that currently has various treatments, like disease-modifying therapies, monoclonal antibodies, corticosteroids, and hormonal derivatives. Melatonin has several actions, like regulating circadian rhythms, which are usually used for insomnia. This scoping review aimed to explore the efficacy of melatonin, both as a standalone treatment and in conjunction with other drugs, in the management of MS in animal models.
We searched PubMed, Web of Science, EMBASE, and Google Scholar using (“melatonin” OR “melatonin receptor”) AND (“multiple sclerosis”) AND (“animal”). Animal studies that evaluated the effects of melatonin on the development, progression, and outcomes of MS were included. Human studies and other types of studies like case reports were excluded. We used narrative synthesis for reporting the results.
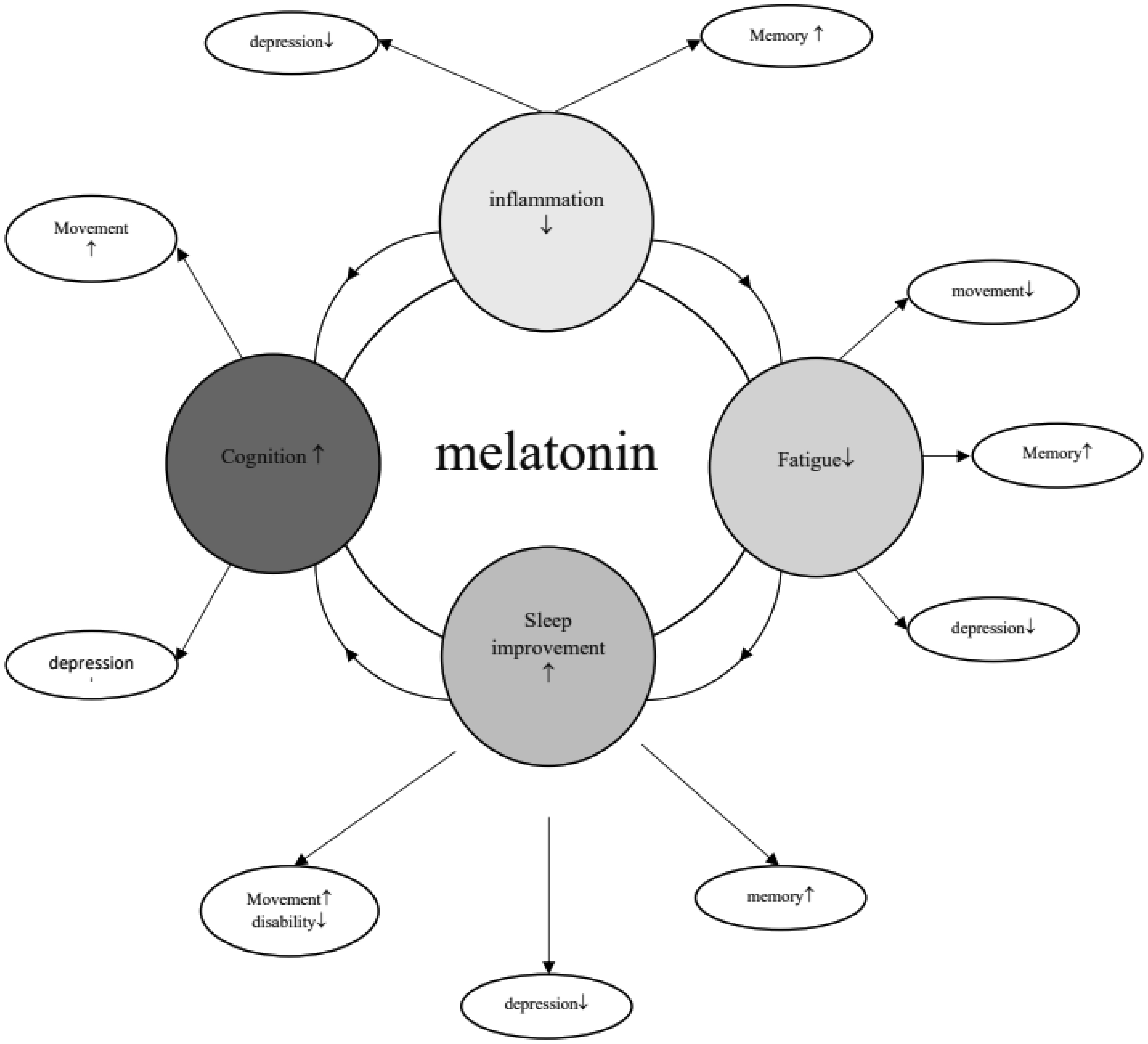
Overall, 21 studies were included, conducted on mice (n = 15) and rats (n = 6). Mostly, studies reported that melatonin led to normal circadian rhythms in animals. Melatonin in doses of both 50 and 100 mg/kg were useful in nociception latency. Melatonin in combination with other drugs like H-89, diisopropylamine dichloroacetate, gibberellins, and IFN-1β improved outcomes, while there was not improved cognition in combination with mesenchymal stem cells. In some tests, male subjects showed significantly better responses. There were controversial results regarding the effects of melatonin on cytokines, but overall, it led to a reduction in proinflammatory cytokines.
Melatonin overall demonstrated favorable outcomes regarding oxidative stress, anti-inflammation, and cytokine levels across various doses. It is recommended conducting systematic reviews and meta-analyses with a larger number of primary studies to provide more comprehensive insights.
Citation: Heliya Bandehagh, Farnaz Gozalpour, Ali Mousavi, Mahdi Hemmati Ghavshough. Effects of melatonin on the management of multiple sclerosis: A scoping review on animal studies[J]. AIMS Medical Science, 2024, 11(2): 137-156. doi: 10.3934/medsci.2024012
Multiple sclerosis (MS), is a debilitating neurological disease that currently has various treatments, like disease-modifying therapies, monoclonal antibodies, corticosteroids, and hormonal derivatives. Melatonin has several actions, like regulating circadian rhythms, which are usually used for insomnia. This scoping review aimed to explore the efficacy of melatonin, both as a standalone treatment and in conjunction with other drugs, in the management of MS in animal models.
We searched PubMed, Web of Science, EMBASE, and Google Scholar using (“melatonin” OR “melatonin receptor”) AND (“multiple sclerosis”) AND (“animal”). Animal studies that evaluated the effects of melatonin on the development, progression, and outcomes of MS were included. Human studies and other types of studies like case reports were excluded. We used narrative synthesis for reporting the results.
Overall, 21 studies were included, conducted on mice (n = 15) and rats (n = 6). Mostly, studies reported that melatonin led to normal circadian rhythms in animals. Melatonin in doses of both 50 and 100 mg/kg were useful in nociception latency. Melatonin in combination with other drugs like H-89, diisopropylamine dichloroacetate, gibberellins, and IFN-1β improved outcomes, while there was not improved cognition in combination with mesenchymal stem cells. In some tests, male subjects showed significantly better responses. There were controversial results regarding the effects of melatonin on cytokines, but overall, it led to a reduction in proinflammatory cytokines.
Melatonin overall demonstrated favorable outcomes regarding oxidative stress, anti-inflammation, and cytokine levels across various doses. It is recommended conducting systematic reviews and meta-analyses with a larger number of primary studies to provide more comprehensive insights.

| [1] |
Jakimovski D, Bittner S, Zivadinov R, et al. (2023) Multiple sclerosis. Lancet 403: 183-202. https://doi.org/10.1016/S0140-6736(23)01473-3 
|
| [2] |
Thompson AJ, Baranzini SE, Geurts J, et al. (2018) Multiple sclerosis. Lancet 391: 1622-1636. https://doi.org/10.1016/S0140-6736(18)30481-1
|
| [3] |
McGinley MP, Goldschmidt CH, Rae-Grant AD (2021) Diagnosis and treatment of multiple sclerosis: a review. JAMA 325: 765-779. https://doi.org/10.1001/jama.2020.26858
|
| [4] |
Hauser SL, Cree BAC (2020) Treatment of multiple sclerosis: a review. Am J Med 133: 1380-1390.e2. https://doi.org/10.1016/j.amjmed.2020.05.049
|
| [5] |
Qian Z, Li Y, Guan Z, et al. (2023) Global, regional, and national burden of multiple sclerosis from 1990 to 2019: findings of global burden of disease study 2019. Front Public Health 11: 1073278. https://doi.org/10.3389/fpubh.2023.1073278
|
| [6] |
Català-Senent JF, Andreu Z, Hidalgo MR, et al. (2023) A deep transcriptome meta-analysis reveals sex differences in multiple sclerosis. Neurobiol Dis 181: 106113. https://doi.org/10.1016/j.nbd.2023.106113
|
| [7] |
Cardinali DP (2019) Melatonin: clinical perspectives in neurodegeneration. Front Endocrinol 10: 480. https://doi.org/10.3389/fendo.2019.00480
|
| [8] |
Gunata M, Parlakpinar H, Acet HA (2020) Melatonin: a review of its potential functions and effects on neurological diseases. Rev Neurol 176: 148-165. https://doi.org/10.1016/j.neurol.2019.07.025
|
| [9] |
Hardeland R (2018) Melatonin and inflammation—story of a double-edged blade. J Pineal Res 65: e12525. https://doi.org/10.1111/jpi.12525
|
| [10] |
Loloei S, Sepidarkish M, Heydarian A, et al. (2019) The effect of melatonin supplementation on lipid profile and anthropometric indices: a systematic review and meta-analysis of clinical trials. Diabetes Metab Syndr 13: 1901-1910. https://doi.org/10.1016/j.dsx.2019.04.043
|
| [11] |
Razmaray H, Nasiri E, Vakilipour P, et al. (2024) The effects of melatonin supplementation on neurobehavioral outcomes and clinical severity in rodent models of multiple sclerosis; a systematic review and meta-analysis. Inflammopharmacology 32: 927-944. https://doi.org/10.1007/s10787-023-01414-7
|
| [12] |
Morsali S, Sabahi Z, Kakaei J, et al. (2023) Clinical efficacy and safety of melatonin supplementation in multiple sclerosis: a systematic review. Inflammopharmacology 31: 2213-2220. https://doi.org/10.1007/s10787-023-01271-4
|
| [13] |
Skarlis C, Anagnostouli M (2020) The role of melatonin in multiple sclerosis. Neurol Sci 41: 769-781. https://doi.org/10.1007/s10072-019-04137-2
|
| [14] |
Muñoz-Jurado A, Escribano BM, Caballero-Villarraso J, et al. (2022) Melatonin and multiple sclerosis: antioxidant, anti-inflammatory and immunomodulator mechanism of action. Inflammopharmacology 30: 1569-1596. https://doi.org/10.1007/s10787-022-01011-0
|
| [15] |
Tricco AC, Lillie E, Zarin W, et al. (2018) PRISMA Extension for Scoping Reviews (PRISMA-ScR): checklist and explanation. Ann Intern Med 169: 467-473. https://doi.org/10.7326/M18-0850
|
| [16] |
Abo Taleb HA, Alghamdi BS (2020) Neuroprotective effects of melatonin during demyelination and remyelination stages in a mouse model of multiple sclerosis. J Mol Neurosci 70: 386-402. https://doi.org/10.1007/s12031-019-01425-6
|
| [17] |
Wen J, Ariyannur PS, Ribeiro R, et al. (2016) Efficacy of N-acetylserotonin and melatonin in the EAE model of multiple sclerosis. J Neuroimmune Pharmacol 11: 763-773. https://doi.org/10.1007/s11481-016-9702-9
|
| [18] |
Escribano BM, Muñoz-Jurado A, Caballero-Villarraso J, et al. (2022) Protective effects of melatonin on changes occurring in the experimental autoimmune encephalomyelitis model of multiple sclerosis. Mult Scler Relat Disord 58: 103520. https://doi.org/10.1016/j.msard.2022.103520
|
| [19] |
Ghareghani M, Dokoohaki S, Ghanbari A, et al. (2017) Melatonin exacerbates acute experimental autoimmune encephalomyelitis by enhancing the serum levels of lactate: a potential biomarker of multiple sclerosis progression. Clin Exp Pharmacol Physiol 44: 52-61. https://doi.org/10.1111/1440-1681.12678
|
| [20] |
Ortíz GG, Briones-Torres AL, Benitez-King G, et al. (2022) Beneficial effect of melatonin alone or in combination with glatiramer acetate and interferon β-1b on experimental autoimmune encephalomyelitis. Molecules 27: 4217. https://doi.org/10.3390/molecules27134217
|
| [21] |
Ghareghani M, Farhadi Z, Rivest S, et al. (2022) PDK4 inhibition ameliorates melatonin therapy by modulating cerebral metabolism and remyelination in an EAE Demyelinating mouse model of multiple sclerosis. Front Immunol 13: 862316. https://doi.org/10.3389/fimmu.2022.862316
|
| [22] |
Ghareghani M, Pons V, Laflamme N, et al. (2023) Inhibiting nighttime melatonin and boosting cortisol increase patrolling monocytes, phagocytosis, and myelination in a murine model of multiple sclerosis. Exp Mol Med 55: 215-227. https://doi.org/10.1038/s12276-023-00925-1
|
| [23] | Gharib D, Rashed L, Yousuf A, et al. (2022) Therapeutic effect of microvesicles derived from BM-MSCS transplantation and/or melatonin in cuprizone model of multiple sclerosis: a pharmacodynamic biochemical assay. Egypt J Chem 65: 153-169. |
| [24] |
González EJR, Jirano LJ, Martínez DZG, et al. (2021) A comparative study of melatonin and immunomodulatory therapy with interferon beta and glatiramer acetate in a mouse model of multiple sclerosis. Neurología 36: 262-270. https://doi.org/10.1016/j.nrleng.2018.01.004
|
| [25] |
Ghareghani M, Scavo L, Arnoult D, et al. (2018) Melatonin therapy reduces the risk of osteoporosis and normalizes bone formation in multiple sclerosis. Fundam Clin Pharmacol 32: 181-187. https://doi.org/10.1111/fcp.12337
|
| [26] |
Jand Y, Ghahremani MH, Ghanbari A, et al. (2022) Melatonin ameliorates disease severity in a mouse model of multiple sclerosis by modulating the kynurenine pathway. Sci Rep 12: 15963. https://doi.org/10.1038/s41598-022-20164-0
|
| [27] |
Labunets I, Rodnichenko A, Savosko S, et al. (2023) Reaction of different cell types of the brain on neurotoxin cuprizone and hormone melatonin treatment in young and aging mice. Front Cell Neurosci 17: 1131130. https://doi.org/10.3389/fncel.2023.1131130
|
| [28] |
Álvarez-Sánchez N, Cruz-Chamorro I, López-González A, et al. (2015) Melatonin controls experimental autoimmune encephalomyelitis by altering the T effector/regulatory balance. Brain Behav Immun 50: 101-114. https://doi.org/10.1016/j.bbi.2015.06.021
|
| [29] |
Chen SJ, Huang SH, Chen JW, et al. (2016) Melatonin enhances interleukin-10 expression and suppresses chemotaxis to inhibit inflammation in situ and reduce the severity of experimental autoimmune encephalomyelitis. Int Immunopharmacol 31: 169-177. https://doi.org/10.1016/j.intimp.2015.12.020
|
| [30] |
Long T, Yang Y, Peng L, et al. (2018) Neuroprotective effects of melatonin on experimental allergic encephalomyelitis mice via anti-oxidative stress activity. J Mol Neurosci 64: 233-241. https://doi.org/10.1007/s12031-017-1022-x
|
| [31] |
Ghareghani M, Zibara K, Sadeghi H, et al. (2018) Spasticity treatment ameliorates the efficacy of melatonin therapy in Experimental Autoimmune Encephalomyelitis (EAE) mouse model of multiple sclerosis. Cell Mol Neurobiol 38: 1145-1151. https://doi.org/10.1007/s10571-018-0580-y
|
| [32] |
Dokoohaki S, Ghareghani M, Ghanbari A, et al. (2017) Corticosteroid therapy exacerbates the reduction of melatonin in multiple sclerosis. Steroids 128: 32-36. https://doi.org/10.1016/j.steroids.2017.10.006
|
| [33] |
Alghamdi BS, AboTaleb HA (2020) Melatonin improves memory defects in a mouse model of multiple sclerosis by up-regulating cAMP-response element-binding protein and synapse-associated proteins in the prefrontal cortex. J Integr Neurosci 19: 229-237. https://doi.org/10.31083/j.jin.2020.02.32
|
| [34] |
Sharif R, Aghsami M, Gharghabi M, et al. (2017) Melatonin reverses H-89 induced spatial memory deficit: involvement of oxidative stress and mitochondrial function. Behav Brain Res 316: 115-124. https://doi.org/10.1016/j.bbr.2016.08.040
|
| [35] |
Labunets IF, Utko NA, Toporova OK (2021) Effects of multipotent mesenchymal stromal cells of the human umbilical cord and their combination with melatonin in adult and aging mice with a toxic cuprizone model of demyelination. Adv Gerontol 11: 173-180. https://doi.org/10.1134/S2079057021020077
|
| [36] |
Vakilzadeh G, Khodagholi F, Ghadiri T, et al. (2016) The effect of melatonin on behavioral, molecular, and histopathological changes in cuprizone model of demyelination. Mol Neurobiol 53: 4675-4684. https://doi.org/10.1007/s12035-015-9404-y
|
| [37] |
Pivovarova-Ramich O, Zimmermann HG, Paul F (2023) Multiple sclerosis and circadian rhythms: can diet act as a treatment?. Acta Physiol 237: e13939. https://doi.org/10.1111/apha.13939
|
| [38] |
Ohl K, Tenbrock K, Kipp M (2016) Oxidative stress in multiple sclerosis: central and peripheral mode of action. Exp Neurol 277: 58-67. https://doi.org/10.1016/j.expneurol.2015.11.010
|
| [39] |
Pereira GM, Soares NM, Souza AR, et al. (2018) Basal cortisol levels and the relationship with clinical symptoms in multiple sclerosis: a systematic review. Arq Neuropsiquiatr 76: 622-634. https://doi.org/10.1590/0004-282X20180091
|
| [40] |
Cohen JA (2013) Mesenchymal stem cell transplantation in multiple sclerosis. J Neurol Sci 333: 43-49. https://doi.org/10.1016/j.jns.2012.12.009
|
| [41] |
Hu C, Li L (2019) Melatonin plays critical role in mesenchymal stem cell-based regenerative medicine in vitro and in vivo. Stem Cell Res Ther 10: 13. https://doi.org/10.1186/s13287-018-1114-8
|
| [42] |
Farhadi N, Oryan S, Nabiuni M (2014) Serum levels of melatonin and cytokines in multiple sclerosis. Biomed J 37: 90-92. https://doi.org/10.4103/2319-4170.125885
|

Figures(2) / Tables(2)

Heliya Bandehagh, Farnaz Gozalpour, Ali Mousavi, Mahdi Hemmati Ghavshough. Effects of melatonin on the management of multiple sclerosis: A scoping review on animal studies[J]. AIMS Medical Science, 2024, 11(2): 137-156. doi: 10.3934/medsci.2024012







 DownLoad:
DownLoad: