Traditional compartmental models of epidemic transmission often predict an initial phase of exponential growth, assuming uniform susceptibility and interaction within the population. However, empirical outbreak data frequently show early stages of sub-exponential growth in case incidences, challenging these assumptions and indicating that traditional models may not fully encompass the complexity of epidemic dynamics. This discrepancy has been addressed through models that incorporate early behavioral changes or spatial constraints within contact networks. In this paper, we propose the concept of "frailty", which represents the variability in individual susceptibility and transmission, as a more accurate approach to understanding epidemic growth. This concept shifts our understanding from a purely exponential model to a more nuanced, generalized model, depending on the level of heterogeneity captured by the frailty parameter. By incorporating this type of heterogeneity, often overlooked in traditional models, we present a novel mathematical framework. This framework enhances our understanding of how individual differences affect key epidemic metrics, including reproduction numbers, epidemic size, likelihood of stochastic extinction, impact of public health interventions, and accuracy of disease forecasts. By accounting for individual heterogeneity, our approach suggests that a more complex and detailed understanding of disease spread is necessary to accurately predict and manage outbreaks.
Citation: Ping Yan, Gerardo Chowell. Modeling sub-exponential epidemic growth dynamics through unobserved individual heterogeneity: a frailty model approach[J]. Mathematical Biosciences and Engineering, 2024, 21(10): 7278-7296. doi: 10.3934/mbe.2024321
Traditional compartmental models of epidemic transmission often predict an initial phase of exponential growth, assuming uniform susceptibility and interaction within the population. However, empirical outbreak data frequently show early stages of sub-exponential growth in case incidences, challenging these assumptions and indicating that traditional models may not fully encompass the complexity of epidemic dynamics. This discrepancy has been addressed through models that incorporate early behavioral changes or spatial constraints within contact networks. In this paper, we propose the concept of "frailty", which represents the variability in individual susceptibility and transmission, as a more accurate approach to understanding epidemic growth. This concept shifts our understanding from a purely exponential model to a more nuanced, generalized model, depending on the level of heterogeneity captured by the frailty parameter. By incorporating this type of heterogeneity, often overlooked in traditional models, we present a novel mathematical framework. This framework enhances our understanding of how individual differences affect key epidemic metrics, including reproduction numbers, epidemic size, likelihood of stochastic extinction, impact of public health interventions, and accuracy of disease forecasts. By accounting for individual heterogeneity, our approach suggests that a more complex and detailed understanding of disease spread is necessary to accurately predict and manage outbreaks.

| [1] |
L. Meyers, Contact network epidemiology: Bond percolation applied to infectious disease prediction and control, Bull. Am. Math. Soc., 44 (2007), 63–86. https://doi.org/10.1090/S0273-0979-06-01148-7 doi: 10.1090/S0273-0979-06-01148-7

|
| [2] |
J. O. Lloyd-Smith, S. J. Schreiber, P. E. Kopp, W. M. Getz, Superspreading and the effect of individual variation on disease emergence, Nature, 438 (2005), 355–359. https://doi.org/10.1038/nature04153 doi: 10.1038/nature04153
|
| [3] | L Danon, A. P. Ford, T. House, C. P. Jewell, M. J. Keeling, G. O. Roberts, et al., Networks and the epidemiology of infectious disease, Interdiscip. Perspect. Infect. Dis., 2011 (2011), 284909. |
| [4] | M. J. Keeling, K. T. Eames, Networks and epidemic models, J. R. Soc. Int., 2 (2005), 295–307. |
| [5] | S. Towers, Z. Feng, Social contact patterns and control strategies for influenza in the elderly, Math. Biosci., 204 (2012), 241–249. |
| [6] |
G. Chowell, L. Sattenspiel, S. Bansal, C. Viboud, Mathematical models to characterize early epidemic growth: A Review, Phys. Life Rev., 18 (2016), 66–97. https://doi.org/10.1016/j.plrev.2016.07.005 doi: 10.1016/j.plrev.2016.07.005
|
| [7] | O. Diekmann, J. Heesterbeek, Mathematical Epidemiology of Infectious Diseases: Model Building, Analysis and Interpretation, Wiley, 2000. |
| [8] |
E. Kenah, M. Lipsitch, J. M. Robins, Generation interval contraction and epidemic data analysis, Math. Biosci., 213 (2008), 71–79. https://doi.org/10.1016/j.mbs.2008.02.007 doi: 10.1016/j.mbs.2008.02.007
|
| [9] |
R. Pastor-Satorras, C. Castellano, P. Van Mieghem, A. Vespignani, Epidemic processes in complex networks, Rev. Mod. Phys., 87 (2015), 925–979. https://doi.org/10.1103/RevModPhys.87.925 doi: 10.1103/RevModPhys.87.925
|
| [10] |
M. J. Keeling, The effects of local spatial structure on epidemiological invasions, Proc. R. Soc. London Ser. B Biol. Sci., 266 (1999), 859–867. https://doi.org/10.1098/rspb.1999.0716 doi: 10.1098/rspb.1999.0716
|
| [11] |
M. Barthélemy, A. Barrat, R. Pastor-Satorras, A. Vespignani, Dynamical patterns of epidemic outbreaks in complex heterogeneous networks, J. Theor. Biol., 235 (2005), 275–288. https://doi.org/10.1016/j.jtbi.2005.01.011 doi: 10.1016/j.jtbi.2005.01.011
|
| [12] |
Y. Wang, Z. Wei, J. Cao, Epidemic dynamics of influenza-like diseases spreading in complex networks, Nonlinear Dyn., 101 (2020), 1801–1820. https://doi.org/10.1007/s11071-020-05867-1 doi: 10.1007/s11071-020-05867-1
|
| [13] | P. Ji, J. Ye, Y. Mu, W. Lin, Y. Tian, C. Hens, et al., Signal propagation in complex networks, Phys. Rep., 1017 (2023), 1–96. |
| [14] |
O. Artime, M. Grassia, M. De Domenico, J. P. Gleeson, H. A. Makse, G. Mangioni, Robustness and resilience of complex networks, Nat. Rev. Phys., 6 (2024), 114–131. https://doi.org/10.1038/s42254-023-00676-y doi: 10.1038/s42254-023-00676-y
|
| [15] |
C. Viboud, L. Simonsen, G. Chowell, A generalized-growth model to characterize the early ascending phase of infectious disease outbreaks, Epidemics, 15 (2016), 27–37. https://doi.org/10.1016/j.epidem.2016.01.002 doi: 10.1016/j.epidem.2016.01.002
|
| [16] |
R. Bürger, G. Chowell, L. Y. Lara-Díaz, Measuring differences between phenomenological growth models applied to epidemiology, Math. Biosci., 334 (2021), 108558. https://doi.org/10.1016/j.mbs.2021.108558 doi: 10.1016/j.mbs.2021.108558
|
| [17] | A. Smirnova, L. deCamp, G. Chowell, Forecasting epidemics through nonparametric estimation of time-dependent transmission rates using the SEIR model, Bull. Math. Biol., 81 (2019), 4343–4365. |
| [18] | D. Shanafelt, G. Jones, M. Lima, C. Perrings, G. Chowell, Forecasting the foot-and-mouth disease epidemic in the United Kingdom, EcoHealth, forthcoming. |
| [19] |
B. Pell, Y. Kuang, C. Viboud, G. Chowell, Using phenomenological models for forecasting the 2015 Ebola challenge, Epidemics, 22 (2018), 62–70. https://doi.org/10.1016/j.epidem.2016.11.002 doi: 10.1016/j.epidem.2016.11.002
|
| [20] | G. Chowell, D. Hincapie-Palacio, J. Ospina, B. Pell, A. Tariq, S. Dahal, et al., Using phenomenological models to characterize transmissibility and forecast patterns and final burden of Zika epidemics, PLOS Curr., 8 (2016). |
| [21] |
G. Chowell, C. Viboud, L. Simonsen, S. Moghadas, Characterizing the reproduction number of epidemics with early sub-exponential growth dynamics, J. R. Soc. Interface, 13 (2016), 20160659. https://doi.org/10.1098/rsif.2016.0659 doi: 10.1098/rsif.2016.0659
|
| [22] | G. Chowell, C. Viboud, Is it growing exponentially fast?–Impact of assuming exponential growth for characterizing and forecasting epidemics with initial near-exponential growth dynamics, Infect. Dis. Model., 1 (2016), 71–78. |
| [23] | P. Yan, G. Chowell, Quantitative methods for investigating infectious disease outbreaks, Cham, Switzerland, Springer, 2019. |
| [24] | N. T. J. Bailey, The Mathematical Theory of Infectious Diseases and Its Applications, Second Edition, The Griffin & Company Ltd, 1975. |
| [25] | F. Brauer, C. Castillo-Chávez, Mathematical Models in Population Biology and Epidemiology, Springer, 2001. |
| [26] | P. Erdös, A. Rényi, On the evolution of random graphs, Bull. Inst. Int. Stat., 38 (1961), 343–347. |
| [27] | A. W. Marshall, I. Olkin, Life Distributions: Structure of Nonparametric, Semiparametric and Parametric Families, Springer, 2007. |
| [28] |
A. S. Novozhilov, On the spread of epidemics in a closed heterogeneous population, Math. Biosci., 215 (2008), 177–185. https://doi.org/10.1182/asheducation-2008.1.177 doi: 10.1182/asheducation-2008.1.177
|
| [29] |
C. Rose, A. J. Medford, C. F. Goldsmith, T. Vegge, J. S. Weitz, A. A. Peterson, Heterogeneity in susceptibility dictates the order of epidemic models, J. Theor. Biol., 528 (2021), 110839. https://doi.org/10.1016/j.jtbi.2021.110839 doi: 10.1016/j.jtbi.2021.110839
|
| [30] |
B. Klar, A note on the $L$-class of life distributions, J. Appl. Prob., 39 (2002), 11–19. https://doi.org/10.1239/jap/1019737984 doi: 10.1239/jap/1019737984
|
| [31] | J. D. Kalbfleisch, R. L. Prentice, Statistical Analysis for Failure Time Data, Second Edition, Wiley & Sons, New York, 2002. |
| [32] | N. L. Komarova, L. M. Schang, D. Wodarz, Patterns of the COVID-19 pandemic spread around the world: exponential versus power laws, J. R. Soc. Interface, 17 (2020), 20200518. |
| [33] |
D. G. Xenikos, A. Asimakopoulos, Power-law growth of the COVID-19 fatality incidents in Europe, Infect. Dis. Modell., 6 (2021), 743–750. https://doi.org/10.1016/j.idm.2021.05.001 doi: 10.1016/j.idm.2021.05.001
|
| [34] | A. Tariq, E. A. Undurraga, C. C. Laborde, K. Vogt-Geisse, R. Luo, R. Rothenberg, et al., Transmission dynamics and control of COVID-19 in Chile, March-October, 2020. PLoS Neglected Trop. Dis., 15 (2021), e0009070. https://doi.org/10.1371/journal.pntd.0009070 |
| [35] |
G. Chowell, S. Dahal, A. Tariq, K. Roosa, J. M. Hyman, R. Luo, An ensemble n-sub-epidemic modeling framework for short-term forecasting epidemic trajectories: Application to the COVID-19 pandemic in the USA, PLoS Comput. Biol., 18 (2022), e1010602. https://doi.org/10.1371/journal.ppat.1010602 doi: 10.1371/journal.ppat.1010602
|
| [36] | G. Chowell, R. Rothenberg, K. Roosa, A. Tariq, J. M. Hyman, R. Luo, Sub-epidemic model forecasts during the first wave of the COVID-19 pandemic in the USA and European hotspots, in Mathematics of Public Health: Proceedings of the Seminar on the Mathematical Modelling of COVID-19, (2022), 85–137. |
| [37] |
P. Yan, A frailty model for intervention effectiveness against disease transmission when implemented with unobservable heterogeneity, Math. Biosci. Eng., 15 (2018), 275–298. https://doi.org/10.3934/mbe.2018012 doi: 10.3934/mbe.2018012
|
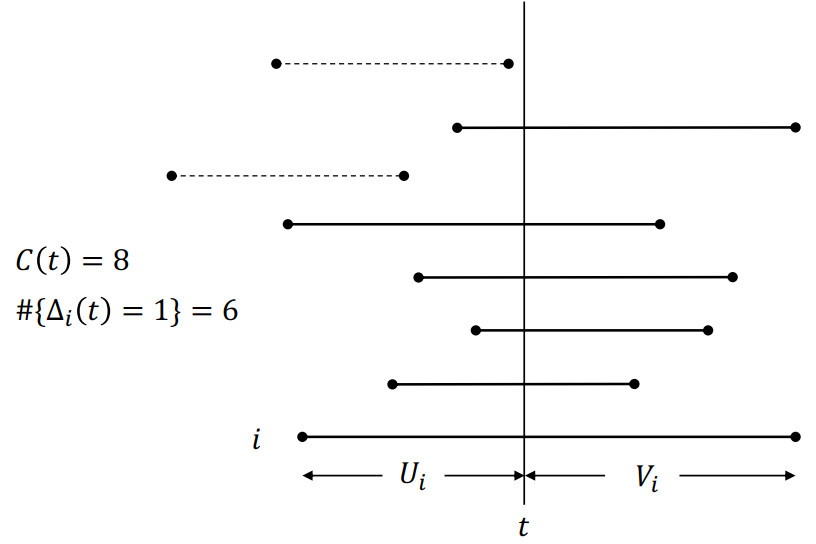


Figures(3)

Ping Yan, Gerardo Chowell. Modeling sub-exponential epidemic growth dynamics through unobserved individual heterogeneity: a frailty model approach[J]. Mathematical Biosciences and Engineering, 2024, 21(10): 7278-7296. doi: 10.3934/mbe.2024321







 DownLoad:
DownLoad: