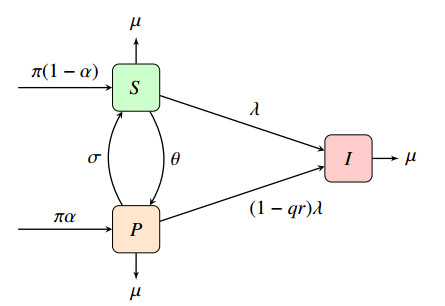
A model with both casual and long-term partnerships is considered with respect to the impact of a pre-exposure prophylaxis (PrEP) on the spread of HIV. We consider the effect of the effectiveness of PrEP, the rate that susceptible individuals choose to take PrEP, and compliance with the daily dose of the pre-exposure prophylaxis. The rate of infection in long-term partnerships is computed using a linearized expected value as a means for including the nonlocal effects of long-term partnerships while maintaining computational feasibility. The reproduction numbers for models with casual partnerships, long-term partnerships, and a combination of both are analytically computed and global stability of both disease-free and endemic equilibria is shown. Sensitivity and PRCC analysis results suggest that increasing the compliance among the current PrEP users is a more effective strategy in the fight against the HIV epidemic than increased coverage with poor compliance. Furthermore, an analysis of the reproduction number shows that models with either casual or monogamous long-term partnerships can reach the desired $ R_0 < 1 $ threshold for high enough levels of compliance and uptake, however, a model with both casual and monogamous long-term partnerships will require additional interventions. Methods highlighted in this manuscript are applicable to other incurable diseases or diseases with imperfect vaccines effected by long-term partnerships.
Citation: S. J. Gutowska, K. A. Hoffman, K. F. Gurski. The effect of PrEP uptake and adherence on the spread of HIV in the presence of casual and long-term partnerships[J]. Mathematical Biosciences and Engineering, 2022, 19(12): 11903-11934. doi: 10.3934/mbe.2022555
A model with both casual and long-term partnerships is considered with respect to the impact of a pre-exposure prophylaxis (PrEP) on the spread of HIV. We consider the effect of the effectiveness of PrEP, the rate that susceptible individuals choose to take PrEP, and compliance with the daily dose of the pre-exposure prophylaxis. The rate of infection in long-term partnerships is computed using a linearized expected value as a means for including the nonlocal effects of long-term partnerships while maintaining computational feasibility. The reproduction numbers for models with casual partnerships, long-term partnerships, and a combination of both are analytically computed and global stability of both disease-free and endemic equilibria is shown. Sensitivity and PRCC analysis results suggest that increasing the compliance among the current PrEP users is a more effective strategy in the fight against the HIV epidemic than increased coverage with poor compliance. Furthermore, an analysis of the reproduction number shows that models with either casual or monogamous long-term partnerships can reach the desired $ R_0 < 1 $ threshold for high enough levels of compliance and uptake, however, a model with both casual and monogamous long-term partnerships will require additional interventions. Methods highlighted in this manuscript are applicable to other incurable diseases or diseases with imperfect vaccines effected by long-term partnerships.

| [1] | CDC, HIV in the United States and dependent areas, 2019. Available from: https://www.cdc.gov/hiv/statistics/overview/ataglance.html. |
| [2] | B. T. Foley, B. T. M. Korber, T. K. Leitner, C. Apetrei, B. Hahn, I. Mizrachi, et al., HIV Sequence Compendium 2018, United States: N. p., 2018. https://doi.org/10.2172/1458915 |
| [3] |
A. S. Fauci, R. R. Redfield, G. Sigounas, M. D. Weahkee, B. P. Giroir, Ending the HIV epidemic: a plan for the United States, JAMA, 321 (2019), 844–845. https://doi.org/10.1001/jama.2019.1343 doi: 10.1001/jama.2019.1343

|
| [4] | CDC, Preexposure prophylaxis for the prevention of HIV infection in the United States - 2017 update: a clinical practice guideline, 2018. Available from: https://www.cdc.gov/hiv/pdf/risk/prep/cdc-hiv-prep-guidelines-2017.pdf. |
| [5] |
J. Franks, Y. Hirsch-Moverman, A. S. Loquere, K. R. Amico, R. M. Grant, B. J. Dye, et al., Sex, PrEP, and stigma: experiences with HIV pre-exposure prophylaxis among New York City MSM participating in the HPTN 067/ADAPT study, AIDS Behav., 22 (2018), 1139–1149. https://doi.org/10.1007/s10461-017-1964-6 doi: 10.1007/s10461-017-1964-6
|
| [6] |
S. McCormack, D. T. Dunn, M. Desai, D. I. Dolling, M. Gafos, R. Gilson, et al., Pre-exposure prophylaxis to prevent the acquisition of HIV-1 infection (PROUD): effectiveness results from the pilot phase of a pragmatic open-label randomised trial, Lancet, 387 (2016), 53–60. https://doi.org/10.1016/S0140-6736(15)00056-2 doi: 10.1016/S0140-6736(15)00056-2
|
| [7] |
J. M. Molina, C. Capitant, B. Spire, G. Pialoux, L. Cotte, I. Charreau, et al., On-demand preexposure prophylaxis in men at high risk for HIV-1 infection, N. Engl. J. Med., 373 (2015), 2237–2246. https://doi.org/10.1056/NEJMoa1506273 doi: 10.1056/NEJMoa1506273
|
| [8] |
D. K. Smith, M. V. Handel, R. J. Wolitski, J. E. Stryker, H. I. Hall, J. Prejean, et al., Vital signs: estimated percentages and numbers of adults with indications for preexposure prophylaxis to prevent HIV acquisition – United States, 2015, Morb. Mortal. Wkly. Rep., 64 (2015), 1291–1295. https://doi.org/10.15585/mmwr.mm6446a4 doi: 10.15585/mmwr.mm6446a4
|
| [9] |
S. B. Kim, M. Yoon, N. S. Ku, M. H. Kim, J. E. Song, J. Y. Ahn, et al., Mathematical modeling of HIV prevention measures including pre-exposure prophylaxis on HIV incidence in South Korea, PLoS One, 9 (2014), e90080. https://doi.org/10.1371/journal.pone.0090080 doi: 10.1371/journal.pone.0090080
|
| [10] |
N. Punyacharoensin, W. J. Edmunds, D. De Angelis, V. Delpech, G. Hart, J. Elford, et al., Modelling the HIV epidemic among MSM in the United Kingdom: quantifying the contributions to HIV transmission to better inform prevention initiatives, AIDS, 29 (2015), 339–349. https://doi.org/10.1097/QAD.0000000000000525 doi: 10.1097/QAD.0000000000000525
|
| [11] | N. Punyacharoensin, W. J. Edmunds, D. De Angelis, V. Delpech, G. Hart, J. Elford, et al., Effect of pre-exposure prophylaxis and combination HIV prevention for men who have sex with men in the UK: a mathematical modelling study, Lancet HIV, 3 (2016), E94–E104. https://doi.org/10.1016/S2352-3018(15)00056-9 |
| [12] |
J. Li, L. Peng, S. Gilmour, J. Gu, Y. Ruan, H. Zou, et al., A mathematical model of biomedical interventions for HIV prevention among men who have sex with men in China, BMC Infect. Dis., 18 (2018). https://doi.org/10.1186/s12879-018-3516-8 doi: 10.1186/s12879-018-3516-8
|
| [13] |
B. Steinegger, I. Iacopini, A. Teixeira, A. Bracci, P. Casanova-Ferrer, A. Antonioni, et al., Non-selective distribution of infectious disease prevention may outperform risk-based targeting, Nat. Commun., 13 (2022), 3028. https://doi.org/10.1038/s41467-022-30639-3 doi: 10.1038/s41467-022-30639-3
|
| [14] |
S. M. Jenness, S. M. Goodreau, E. Rosenberg, E. N. Beylerian, K. W. Hoover, D. K. Smith, et al., Impact of the Centers for Disease Control's HIV preexposure prophylaxis guidelines for men who have sex with men in the United States, J. Infect. Dis., 214 (2016), 1800–1807. https://doi.org/10.1093/infdis/jiw223 doi: 10.1093/infdis/jiw223
|
| [15] |
L. Simpson, A. B. Gumel, Mathematical assessment of the role of pre-exposure prophylaxis on HIV transmission dynamics, Appl. Math. Comput., 293 (2017), 168–193. https://doi.org/10.1016/j.amc.2016.07.0433 doi: 10.1016/j.amc.2016.07.0433
|
| [16] |
C. J. Silva, D. F. Torres, Modeling and optimal control of HIV/AIDS prevention through PrEP, Discrete Contin. Dyn. Syst. - S, 11 (2018), 119–140. https://doi.org/10.3934/dcdss.2018008 doi: 10.3934/dcdss.2018008
|
| [17] |
D. Hansson, S. Strömdahl, K. Y. Leung, T. Britton, Introducing pre-exposure prophylaxis to prevent HIV acquisition among men who have sex with men in Sweden: Insights from a mathematical pair formation model, BMJ Open, 10 (2020), e033852. https://doi.org/10.1136/bmjopen-2019-033852 doi: 10.1136/bmjopen-2019-033852
|
| [18] | NIDA, What is HAART? 2020. Available from: https://nida.nih.gov/publications/research-reports/hivaids/what-haart. |
| [19] | K. F. Gurski, K. A. Hoffman, S. J. Gutowska, B. Batista, Modeling HIV and HSV-2 using partnership models, in Contemporary Research in Mathematical Biology. https://doi.org/10.1142/12639 |
| [20] |
K. F. Gurski, A sexually transmitted infection model with long-term partnerships in homogeneous and heterogenous populations, Infect. Dis. Modell., 4 (2019), 142–160. https://doi.org/10.1016/j.idm.2019.05.002 doi: 10.1016/j.idm.2019.05.002
|
| [21] |
K. P. Hadeler, Pair formation, J. Math. Biol., 64 (2012), 613–645. https://doi.org/10.1007/s00285-011-0454-0 doi: 10.1007/s00285-011-0454-0
|
| [22] |
M. Kretzschmar, J. C. Heijne, Pair formation models for sexually transmitted infections: a primer, Infect. Dis. Modell., 2 (2017), 368–378. https://doi.org/10.1016/j.idm.2017.07.002 doi: 10.1016/j.idm.2017.07.002
|
| [23] |
K. Y. Leung, M. Kretzschmar, O. Diekmann, SI infection on a dynamic partnership network: characterization of $R_0$, J. Math. Biol., 71 (2015), 1–56. https://doi.org/10.1007/s00285-014-0808-5 doi: 10.1007/s00285-014-0808-5
|
| [24] | C. Castillo-Chavez, Z. Feng, W. Huang, On the computation of $R_0$ and its role on global stability, in Mathematical Approaches for Emerging and Re-emerging Infectious Diseases: An Introduction, 125 (2002), 31–65. |
| [25] |
E. H. Elbasha, C. N. Podder, A. B. Gumel, Analyzing the dynamics of an SIRS vaccination model with waning natural and vaccine-induced immunity, Nonlinear Anal. Real World Appl., 12 (2011), 2692–2705. https://doi.org/10.1016/j.nonrwa.2011.03.015 doi: 10.1016/j.nonrwa.2011.03.015
|
| [26] |
A. Korobeinikov, G. C. Wake, Lyapunov functions and global stability for SIR, SIRS, and SIS epidemiological models, Appl. Math. Lett., 15 (2002), 955–960. https://doi.org/10.1016/S0893-9659(02)00069-1 doi: 10.1016/S0893-9659(02)00069-1
|
| [27] |
M. Martcheva, Methods for deriving necessary and sufficient conditions for backward bifurcation, J. Biol. Dyn., 13 (2019), 538–566. https://doi.org/10.1080/17513758.2019.1647359 doi: 10.1080/17513758.2019.1647359
|
| [28] |
J. M. Hyman, J. Li, E. A. Stanley, The differential infectivity and staged progression models for the transmission of HIV, Math. Biosci., 155 (1999), 77–109. https://doi.org/10.1016/S0025-5564(98)10057-3 doi: 10.1016/S0025-5564(98)10057-3
|
| [29] |
S. Marino, I. B. Hogue, C. J. Ray, D. E. Kirschner, A methodology for performing global uncertainty and sensitivity analysis in systems biology, J. Theor. Biol., 254 (2008), 178–196. https://doi.org/10.1016/j.jtbi.2008.04.011 doi: 10.1016/j.jtbi.2008.04.011
|
| [30] |
S. M. Blower, H. Dowlatabadi, Sensitivity and uncertainty analysis of complex models of disease transmission: an HIV model, as an example, Int. Stat. Rev., 62 (1994), 229–243. https://doi.org/10.2307/1403510 doi: 10.2307/1403510
|
| [31] |
J. C. Hojilla, D. Vlahov, P. C. Crouch, C. Dawson-Rose, K. Freeborn, A. Carrico, HIV pre-exposure prophylaxis (PrEP) uptake and retention among men who have sex with men in a community-based sexual health clinic, AIDS Behav., 22 (2018), 1096–1099. https://doi.org/10.1007/s10461-017-2009-x doi: 10.1007/s10461-017-2009-x
|
| [32] |
P. A. Chan, L. Mena, R. Patel, C. E. Oldenburg, L. Beauchamps, A. G. Perez-Brumer, et al., Retention in care outcomes for HIV pre-exposure prophylaxis implementation programmes among men who have sex with men in three US cities, J. Int. AIDS Soc., 19 (2016), 20903. https://doi.org/10.7448/IAS.19.1.20903 doi: 10.7448/IAS.19.1.20903
|
| [33] |
C. A. Koss, E. D. Charlebois, J. Ayieko, D. Kwarisiima, J. Kabami, L. B. Balzer, et al., Uptake, engagement, and adherence to pre-exposure prophylaxis offered after population HIV testing in rural Kenya and Uganda: 72-week interim analysis of observational data from the SEARCH study, Lancet HIV, 7 (2020), E249–E261. https://doi.org/10.1016/S2352-3018(19)30433-3 doi: 10.1016/S2352-3018(19)30433-3
|
| [34] |
Z. Baker, M. Javanbakht, S. Mierzwa, C. Pavel, M. Lally, G. Zimet, et al., Predictors of over-reporting HIV pre-exposure prophylaxis (PrEP) adherence among young men who have sex with men (YMSM) in self-reported versus biomarker data, AIDS Behav., 22 (2018), 1174–1183. https://doi.org/10.1007/s10461-017-1958-4 doi: 10.1007/s10461-017-1958-4
|





Figures(6) / Tables(2)

S. J. Gutowska, K. A. Hoffman, K. F. Gurski. The effect of PrEP uptake and adherence on the spread of HIV in the presence of casual and long-term partnerships[J]. Mathematical Biosciences and Engineering, 2022, 19(12): 11903-11934. doi: 10.3934/mbe.2022555










 DownLoad:
DownLoad: