Near-optimization is as sensible and important as optimization for both theory and applications. This paper concerns the near-optimal control of an avian influenza model with saturation on heterogeneous complex networks. Firstly, the basic reproduction number $ \mathcal{R}_{0} $ is defined for the model, which can be used to govern the threshold dynamics of influenza disease. Secondly, the near-optimal control problem was formulated by slaughtering poultry and treating infected humans while keeping the loss and cost to a minimum. Thanks to the maximum condition of the Hamiltonian function and the Ekeland's variational principle, we establish both necessary and sufficient conditions for the near-optimality by several delicate estimates for the state and adjoint processes. Finally, a number of examples presented to illustrate our theoretical results.
Citation: Keguo Ren, Xining Li, Qimin Zhang. Near-optimal control and threshold behavior of an avian influenza model with spatial diffusion on complex networks[J]. Mathematical Biosciences and Engineering, 2021, 18(5): 6452-6483. doi: 10.3934/mbe.2021321
Near-optimization is as sensible and important as optimization for both theory and applications. This paper concerns the near-optimal control of an avian influenza model with saturation on heterogeneous complex networks. Firstly, the basic reproduction number $ \mathcal{R}_{0} $ is defined for the model, which can be used to govern the threshold dynamics of influenza disease. Secondly, the near-optimal control problem was formulated by slaughtering poultry and treating infected humans while keeping the loss and cost to a minimum. Thanks to the maximum condition of the Hamiltonian function and the Ekeland's variational principle, we establish both necessary and sufficient conditions for the near-optimality by several delicate estimates for the state and adjoint processes. Finally, a number of examples presented to illustrate our theoretical results.

| [1] |
E. C. J. Claas, A. D. Osterhaus, R. Van Beek, J. C. De Jong, G. F. Rimmelzwaan, D. A. Senne, et al., Erratum: Human influenza A H5N1 virus related to a highly pathogenic avian influenza virus, Lancet, 351 (1998), 472–477. doi: 10.1016/S0140-6736(97)11212-0

|
| [2] |
W. O. Kermack, A. G. McKendrick, A contribution to the mathematical theory of epidemics, Proc. R. Soc. London, Ser. A, 115(1927), 700–721. doi: 10.1098/rspa.1927.0118
|
| [3] |
W. O. Kermack, A. G. McKendrick, Contributions to the mathematical theory of epidemics. Ⅱ. The problem of endemicity, Proc. R. Soc. London, Ser. A, 138 (1932), 55–83. doi: 10.1098/rspa.1932.0171
|
| [4] |
W. O.Kermack, A. G. McKendrick, Contributions to the mathematical theory of epidemics. Ⅲ. Further studies of the problem of endemicity, Proc. R. Soc. Lond. Ser. A, 141 (1933), 94–122. doi: 10.1098/rspa.1933.0106
|
| [5] |
S. Liu, S. Ruan, X. Zhang, Nonlinear dynamics of avian influenza epidemic models, Math. Biosci., 283 (2017), 118–135. doi: 10.1016/j.mbs.2016.11.014
|
| [6] |
L. Bourouiba, S. Gourley, R. Liu, J. Wu, The interaction of migratory birds and domestic poultry and its role in sustaining avian influenza, SIAM J. Appl. Math., 71 (2011), 487–516. doi: 10.1137/100803110
|
| [7] |
S. Iwami, Y. Takeuchi, X. N. Liu, Avian-human influenza epidemic model, Math. Biosci., 207 (2007), 1–25. doi: 10.1016/j.mbs.2006.08.001
|
| [8] |
N. Tuncer, M. Martcheva, Modeling seasonality in avian influenza H5N1, J. Biol. Syst., 21 (2013), 1340004. doi: 10.1142/S0218339013400044
|
| [9] | X. Hu, Media reports on the impact of bird flu transmission, J. Northwest. Polytech. Univ., 44 (2014), 525–527. |
| [10] |
V. Capasso, G. Serio, A generalization of the Kermack-McKendrick deterministic epidemic model, Math. Biosci., 42 (1978), 43–61. doi: 10.1016/0025-5564(78)90006-8
|
| [11] |
W. M. Liu, S. A. Levin, Y. Iwasa, Influence of nonlinear incidence rates upon the behavior of SIRS epidemiological models, J. Math. Biol., 23 (1986), 187–204. doi: 10.1007/BF00276956
|
| [12] |
D. Xiao, S. Ruan, Global analysis of an epidemic model with nonmonotone incidence rate, Math. Biosci., 208 (2007), 419–429. doi: 10.1016/j.mbs.2006.09.025
|
| [13] | Q. Tang, J. Ge, Z. Lin, An SEI-SI avian-human influenza model with diffusion and nonlocal delay, Appl. Math. Comput., 247 (2014), 753–761. |
| [14] |
C. X. Lei, K. Kwangik, Z. G. Lin, The spreading frontiers of avian-human influenza described by the free boundary, Sci. China, 57 (2014), 971–990. doi: 10.1007/s11425-013-4652-7
|
| [15] |
A. L. Barabasi, R. Albert, Emergence of scaling in random networks, Science, 286 (1999), 509–512. doi: 10.1126/science.286.5439.509
|
| [16] | X. X. Zhan, C. Liu, G. Zhou, Z. K. Zhang, Coupling dynamics of epidemic spreading and information diffusion on complex networks, Appl. Math. Comput., 332 (2018), 437–448. |
| [17] |
P. Van den Driessche, J. Watmough, Reproduction number and sub-threshold endemic equilibria for compartmental models of disease transmission, Math. Biosci., 180 (2002), 29–48. doi: 10.1016/S0025-5564(02)00108-6
|
| [18] |
K. I. Kim, Z. G. Lin, L. Zhang, Avian-human influenza epidemic model with diffusion, Nonlinear Anal., 11 (2010), 313–322. doi: 10.1016/j.nonrwa.2008.11.015
|
| [19] |
G. Dhariwal, A. Jungel, N. Zamponi, Global martingale solutions for a stochastic population cross-diffusion system, Stoch. Process. Their Appl., 129 (2019), 3792–3820. doi: 10.1016/j.spa.2018.11.001
|
| [20] | H. L. Smith, Monotone Dynamical Systems: An Introduction to the Theory of Competitive and Cooperative Systems, American Mathematical Soc., 2008. |
| [21] | R. H. Martin, H. L. Smith, Abstract functional-differential equations and reaction-diffusion systems, Trans. Am. Math. Soc., 321 (1990), 1–44. |
| [22] | R. S. Cantrell, C. Cosner, Spatial Ecology Via Reaction-Diffusion Equations, John Wiley and Sons, Ltd., 2004. |
| [23] | F. H. Clarke, Optimization and nonsmooth analysis, SIAM, Philadelphia, 1990. |
| [24] |
J. Huang, X. Li, G. Wang, Near-optimal control problems for linear forward-backward stochastic systems, Automatica, 46 (2010), 397–404. doi: 10.1016/j.automatica.2009.11.016
|
| [25] |
I. Ekeland, Nonconvex mininization problems, Bull. Am. Math. Soc., 1 (1979), 443–474. doi: 10.1090/S0273-0979-1979-14595-6
|
| [26] |
W. D. Wang, X. Zhao, Basic reproduction numbers for reaction-diffusion epidemic models, SIAM J. Appl. Dyn. Sys., 11 (2012), 1652–1673. doi: 10.1137/120872942
|
| [27] |
H. R. Thieme, Spectral bound and reproduction number for infinite-dimensional population structure and time heterogeneity, SIAM J. Appl. Math., 70 (2009), 188–211. doi: 10.1137/080732870
|
| [28] |
W. Wang, X. Q. Zhao, Basic reproduction numbers for reaction-diffusion epidemic models, SIAM J. Appl. Dyn. Sys., 11 (2012), 1652–1673. doi: 10.1137/120872942
|
| [29] |
L. J. S. Allen, B. M. Bolker, Y. Lou, A. L. Nevai, Asymptotic profiles of the steady states for an SIS epidemic reaction- diffusion model, Discrete Cont. Dyn., 21 (2008), 1. doi: 10.3934/dcds.2008.21.1
|
| [30] |
W. D. Wang, X. Q. Zhao, Spatial invasion threshold of lyme disease, SIAM J. Appl. Math., 75 (2015), 1142–1170. doi: 10.1137/140981769
|
| [31] | X. Mao, Stochastic Differential Equations and Applications, Elsevier, 2007. |
| [32] | J. Yong, X. Zhou, Stochastic Control: Hamiltonian Systems and HJB Equations, Springer, 1999. |
| [33] |
D. Higham, An algorithmic introduction to numerical simulation of stochastic differential equations, SIAM Rev., 43 (2001), 525–546. doi: 10.1137/S0036144500378302
|
| [34] | S. Lenhart, J. T. Workman, Optimal Control Applied to Biological Models, Boca Raton, FL: Chapman and Hall/CRC, 2007. |
| [35] | F. Chen, J. Cui, Cross-species epidemic dynamic model of influenza, in 2016 9th International Congress on Image and Signal Processing, BioMedical Engineering and Informatics (CISP-BMEI), IEEE, (2016), 1567–1572. |
| [36] | China Agricultural Yearbook Editing Committee, China Agriculture Yearbook, 2012. |
| [37] |
S. Liu, S. Ruan, X. Zhang, Nonlinear dynamics of avian influenza epidemic models, Math. Biosci., 283 (2017), 118–135. doi: 10.1016/j.mbs.2016.11.014
|
| [38] |
X. Zhang, Global dynamics of a stochastic avian-human influenza epidemic model with logistic growth for avian population, Nonlinear Dyn., 90 (2017), 2331–2343. doi: 10.1007/s11071-017-3806-5
|
| [39] | H. L. Smith, Monotone Dynamical Systems: An Introduction to the Theory of Competitive and Cooperative Systems, American Mathematical Soc., 2008. |
| [40] |
M. Uzunca, T. Kucukseyhan, H. Yucel, B. Karasozen, Optimal control of convective fitzhughnagumo equation, Comput. Math. Appl., 73 (2017), 2151–2169. doi: 10.1016/j.camwa.2017.02.028
|
| [41] |
R. Buchholz, H. Engel, E. Kammann, Erratum to: on the optimal control of the schlögl-model, Comput. Opy. Appl., 56 (2013), 187–188. doi: 10.1007/s10589-013-9570-7
|
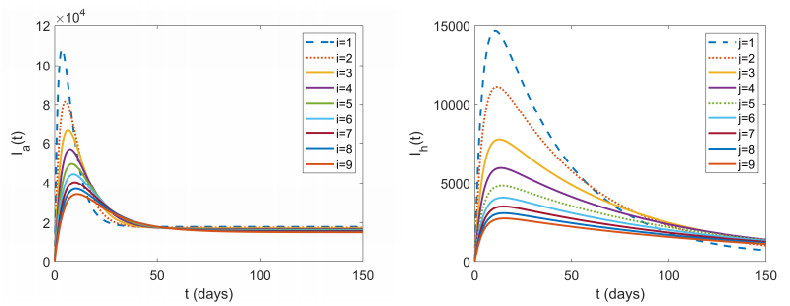
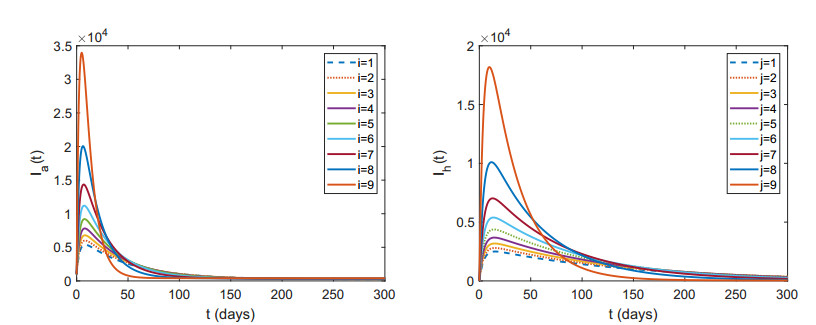
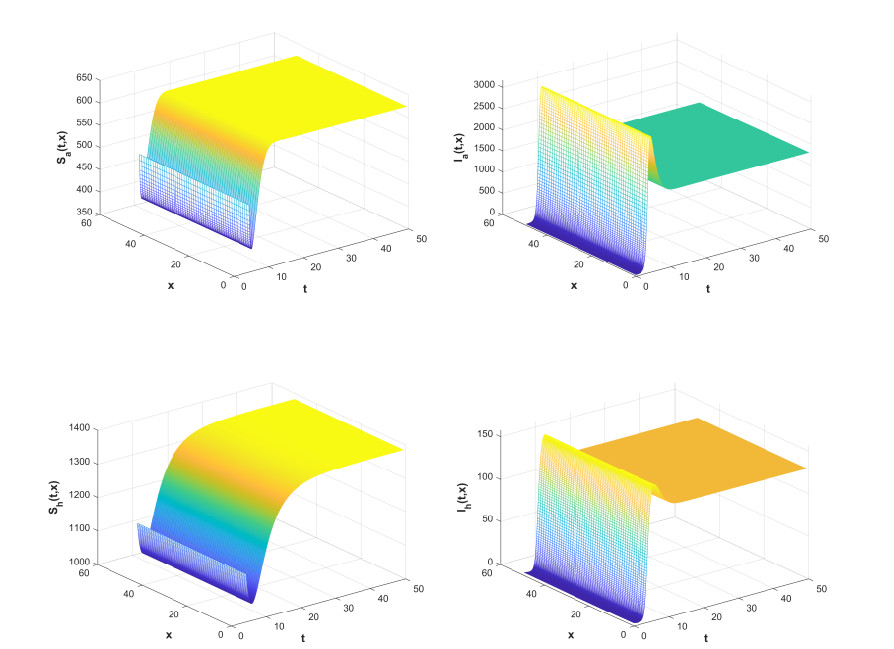
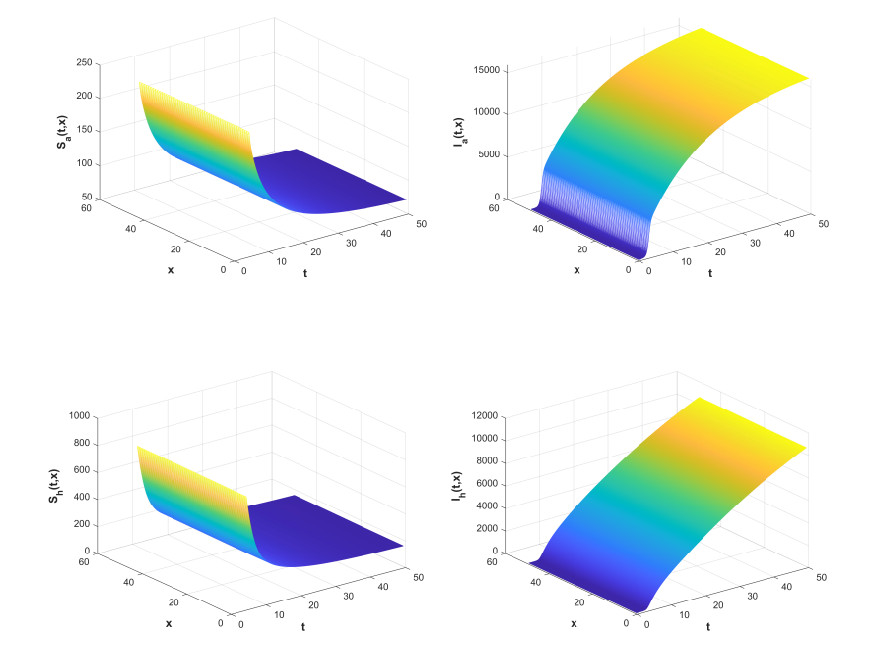
Figures(7) / Tables(2)

Keguo Ren, Xining Li, Qimin Zhang. Near-optimal control and threshold behavior of an avian influenza model with spatial diffusion on complex networks[J]. Mathematical Biosciences and Engineering, 2021, 18(5): 6452-6483. doi: 10.3934/mbe.2021321







 DownLoad:
DownLoad: