Heart diseases are one of the leading causes of death worldwide, and a dysfunction of the cardiac electrical mechanisms is responsible for a significant portion of these deaths. One of these mechanisms, the mechano-electric feedback (MEF), is the electrical response of the heart to local mechanical changes in the environment. This electrical response, in turn, leads to macroscopic changes in heart function. In this paper, we demonstrate that the MEF plays a crucial role in mechanical generation and recovery from arrhythmia which has been observed in experimental studies. To this end, we investigate the cardiac response to a mechanical stimulation using a minimal, multiscale model of the heart which couples the organ level dynamics (left ventricular pressure and volume) and contractile dynamics. By including a mechanical stimulation into the model as a (short, sudden) impulse in the muscle microscale stress, we investigate how the timing, amplitude and duration of the impulse affect the cardiac cycle. In particular, when introduced in the diastolic period of the cardiac cycle, the pulse rate can be stabilised, and ectopic beats and bifurcation can be eliminated, either temporarily or permanently. The stimulation amplitude is a key indicator to this response. We find an optimal value of the impulse amplitude above or below which the impulse maximises the stabilisation. As a result a dysfunction of the MEF can be helped using a mechanical stimulation, by allowing the heart to recover its pumping power. On the other hand, when the mechanical stimulation is introduced towards the end of systole, arrhythmia can be generated.
Citation: Nicholas Pearce, Eun-jin Kim. Modelling the cardiac response to a mechanical stimulation using a low-order model of the heart[J]. Mathematical Biosciences and Engineering, 2021, 18(4): 4871-4893. doi: 10.3934/mbe.2021248
Heart diseases are one of the leading causes of death worldwide, and a dysfunction of the cardiac electrical mechanisms is responsible for a significant portion of these deaths. One of these mechanisms, the mechano-electric feedback (MEF), is the electrical response of the heart to local mechanical changes in the environment. This electrical response, in turn, leads to macroscopic changes in heart function. In this paper, we demonstrate that the MEF plays a crucial role in mechanical generation and recovery from arrhythmia which has been observed in experimental studies. To this end, we investigate the cardiac response to a mechanical stimulation using a minimal, multiscale model of the heart which couples the organ level dynamics (left ventricular pressure and volume) and contractile dynamics. By including a mechanical stimulation into the model as a (short, sudden) impulse in the muscle microscale stress, we investigate how the timing, amplitude and duration of the impulse affect the cardiac cycle. In particular, when introduced in the diastolic period of the cardiac cycle, the pulse rate can be stabilised, and ectopic beats and bifurcation can be eliminated, either temporarily or permanently. The stimulation amplitude is a key indicator to this response. We find an optimal value of the impulse amplitude above or below which the impulse maximises the stabilisation. As a result a dysfunction of the MEF can be helped using a mechanical stimulation, by allowing the heart to recover its pumping power. On the other hand, when the mechanical stimulation is introduced towards the end of systole, arrhythmia can be generated.

| [1] | P. Kohl, P. Hunter, D. Noble, Stretch-induced changes in heart rate and rhythm: clinical observations, experiments and mathematical models, Biophys. Mol. Biol., 71 (1999), 91–138. |
| [2] | T. A, Quinn, P. Kohl, Cardiac Mechano-Electric Coupling: Acute Effects of Mechanical Stimulation on Heart Rate and Rhythm, Physiol Rev., 101 (2021), 37–92. |
| [3] | A. Quarteroni, T. Lassila, S. Rossi, R. Ruiz-Baier, Integrated Heart—Coupling multiscale and multiphysics models for the simulation of the cardiac function, Comput. Methods Appl. Mech. Eng., 314 (2017), 345–407. |
| [4] | Z. Knudsen, A. Holden, J. Brindley, Qualitative modeling of mechano-electrical feedback in a ventricular cell, Bull. Math. Biol., 6 (1997), 115–181. |
| [5] | S. N. Healy, A. D. McCulloch, An ionic model of stretch-activated and stretch-modulated currents in rabbit ventricular myocytes, Europace, 2 (2006), 128–134. |
| [6] | G. Tse, S. T. Wong, V. Tse, Y. T. Lee, H. Y. Lin, J. M. Yeo, Cardiac dynamics: Alternans and arrhythmogenesis, J. Arrhythm, 32 (2016), 411–417. |
| [7] | T. A. Quinn, H. Jin, P. Lee, P. Kohl, Mechanically Induced Ectopy via Stretch-Activated Cation-Nonselective Channels Is Caused by Local Tissue Deformation and Results in Ventricular Fibrillation if Triggered on the Repolarization Wave Edge (Commotio Cordis), Circ Arrhythm Electrophysiol., 8 (2017), e004777. |
| [8] | T. L. Riemer, E. A. Sobie, L. Tung, Stretch-induced changes in arrhythmogenesis and excitability in experimentally based heart cell models, Am. J. Physiol., 275 (1998), 431–442. |
| [9] | P. Kohl, C. Bollensdorff, A. Garny, Effects of mechanosensitive ion channels on ventricular electrophysiology: experimental and theoretical models, Exp Physiol., 91 (2006), 307–328. |
| [10] | M, Zabel, S. Portnoy, M.R. Franz, Effect of sustained load on dispersion of ventricular repolarization and conduction time in the isolated intact rabbit heart, J. Cardiovasc. Electrophysiol., 7 (1996), 9–16. |
| [11] | M. R. Franz, J. Schaefer, M. Schottler, W. A. Seed, M. I. M. Noble, Electrical and mechanical restitution of the human heart at different rates of stimulation, Circ. Res., 53 (1982), 815–822. |
| [12] | N. Westerhof, N. Stergiopulos, M. I. M. Noble, Snapshots of Hemodynamics: An Aid for Clinical Research and Graduate Education, Springer US, 2005. |
| [13] | R. FitzHugh, Impulses and physiological states in theoretical models of nerve membrane, Bull. Math. Biol., 1 (1961), 445-466e. |
| [14] | N. A. Vikulova, L. B. Katsnelson, A. G. Kursanov, O. Solovyova, V. S. Markhasin, Mechano-electric feedback in one-dimensional model of myocardium, J. Math. Biol., 2 (2016), 335–366. |
| [15] | A. Collet, J Bragard, P. C. Dauby, Temperature, geometry, and bifurcations in the numerical modeling of the cardiac mechano-electric feedback, Chaos Interdiscip. J. Nonlinear Sci., 27 (2017), 093924. |
| [16] | A. Gizzi, A. Loppini, R. Ruiz-Baier, A. Ippolito, A. Camassa, A. La Camera, et al., Nonlinear diffusion and thermo-electric coupling in a two-variable model of cardiac action potential, Chaos, 27 (2017), 093919. |
| [17] | Z. Qu, G. Hu, A. Garfinkel, J. N. Weiss, Nonlinear and stochastic dynamics in the heart, Phys. Rep., 543 (2014), 61–162. |
| [18] | C. Franzone, L. F. Pavarino, S. Scacchiet, Effects of mechanical feedback on the stability of cardiac scroll waves: A bidomain electro-mechanical simulation study, Chaos, 27 (2017), 093905. |
| [19] | A. Amar, S. Zlochiver, O. Barnea, Mechano-electric feedback effects in a three-dimensional (3D) model of the contracting cardiac ventricle, PLoS One, 13 (2018), e0191238. |
| [20] | A. Hazim, Y. Belhamadia, S. Dubljevic, Effects of mechano-electrical feedback on the onset of alternans: A computational study, Chaos, 29 (2019), 063126. |
| [21] | M. S. Link, P. J. Wang, N. G. Pandian, S. Bharati, J. E. Udelson, M. Y. Lee, et al., An experimental model of sudden death due to low-energy chest-wall impact (commotio cordis), N. Engl. J. Med., 18 (1998), 1805–1811. |
| [22] | C. Madias, B. J. Maron, A. A. Alsheikh-Ali, M. Rajab, N. A. Estes 3rd, M. S. Link, Precordial thump for cardiac arrest is effective for asystole but not for ventricular fibrillation, Heart Rhythm, 3 (2009), 1495–1500. |
| [23] | W. Li, P. Kohl, N. Trayanova, Myocardial ischemia lowers precordial thump efficacy: an inquiry into mechanisms using three-dimensional simulations, Heart Rhythm, 3 (2006), 179–186. |
| [24] | T. A. Quinn, P. Kohl, Comparing maximum rate and sustainability of pacing by mechanical vs. electrical stimulation in the Langendorff-perfused rabbit heart, Europace, 18 (2016), iv85–iv93. |
| [25] | M. S. Link, P. J. Wang, B. A. VanderBrink, E. Avelar, N. G. Pandian, B. J. Maron, et al., Selective activation of the KATP+ channel is a mechanism by which sudden death is produced by low-energy chest-wall impact (Commotio cordis), Circulation, 100 (1999), 413–418. |
| [26] | B. J. Maron, M. S. Link, P. J. Wang, N. A. Estes 3rd, Clinical profile of commotio cordis: an under appreciated cause of sudden death in the young during sports and other activities, J. Cardiovasc. Electrophysiol., 10 (1999), 114–120. |
| [27] | A. Garny, P. Kohl, Mechanical induction of arrhythmias during ventricular repolarization: modeling cellular mechanisms and their interaction in two dimensions, Ann. N. Y. Acad. Sci., 1015 (2004), 133–143. |
| [28] | W. Li, P. Kohl, N. Trayanova, Induction of ventricular arrhythmias following mechanical impact: A simulation study in 3D, J. Mol. Histol., 35 (2004), 679–686. |
| [29] | E. Kim, M. Capoccia, Synergistic model of cardiac function with a heart assist device, Bioengineering, 7 (2019), 1–16. |
| [30] | K. Sagawa, The ventricular pressure-volume diagram revisited, Circ. Res., 43 (1978), 677–687. |
| [31] | M. A. Simaan, A. Ferreira, S. Chen, J. F. Antaki, D. G. Galati, A dynamical state space representation and performance analysis of feedback-controlled rotary left ventricular assist device, IEEE Trans. Control Syst. Tech., 17 (2019), 15–28. |
| [32] | T. E. Claessens, D. Georgakopoulos, M. Afanasyeva, S. J. Vermeersch, H. D. Millar, N. Stergiopulos, et al., Nonlinear isochrones in murine left ventricular pressure-volume loops: how well does the time-varying elastance concept hold?, Am. J. Physiol. Heart Circ. Physiol., 290 (2006), H1474–H1483. |
| [33] | E. Kim, M. Capoccia, Mechano-electric effect and a heart assist device in the synergistic model of cardiac function, Math. Biosci. Eng., 17 (2020), 5212–5233. |
| [34] | M. R. Franz, Mechano-electrical feedback in ventricular myocardium, Cardiovascular Res., 32 (1996), 263–266. |
| [35] | M. A. Simaan, A. Ferreira, S. Chen, J. F. Antaki and D. G. Galati, A Dynamical State Space Representation and Performance Analysis of a Feedback-Controlled Rotary Left Ventricular Assist Device, IEEE Trans. Control Syst. Technol., 3 (2009), 15–28. |
| [36] | J. Bestel, Modele differentiel de la contraction musculaire controlee: Application au systeme cardio-vasculaire, Ph.D thesis, Universit Paris 9, 2000. |
| [37] | J. Sainte-Marie, D. Chapelle, R. Cimrman, M. Sorine, Modeling and estimation of the cardiac electromechanical activity, Comput. Struct., 84 (2006), 1743–1759. |
| [38] | J. Bestel, F. Clément, M. Sorine, A biomechanical model of muscle contraction, in Medical Image Computing and Computer-Assisted Intervention MICCAI 2001, Springer: Berlin, Germany, (2001), 1159–1161. |
| [39] | P. Krejci, J. Marie, M. Sorine, J. M. Urquiza, Modelling and simulation of an active fibre for cardiac muscle, 2006. Available from: http://www.crm.umontreal.ca/pub/Rapports/3200-3299/3208.pdf. |
| [40] | A. M. Katz, Physiology of the Heart, Lippincott Williams & Wilkins, 2010. |
| [41] | G. Giantesio, A Musesti, D, Riccobelli, A Comparison Between Active Strain and Active Stress in Transversely Isotropic Hyperelastic Materials, J. Elasticity, 137 (2019), 63–82. |
| [42] | A. Gizzi, C. Cherubini, S. Filippi, A. Pandolfi, Theoretical and Numerical Modeling of Nonlinear Electromechanics with applications to Biological Active Media, Communications in Computational Physics, 17 (2015), 93–126. |
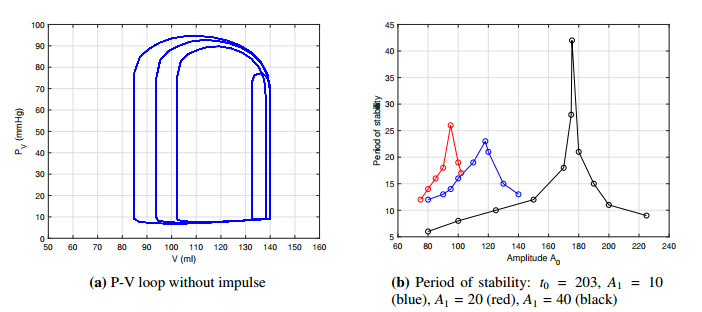
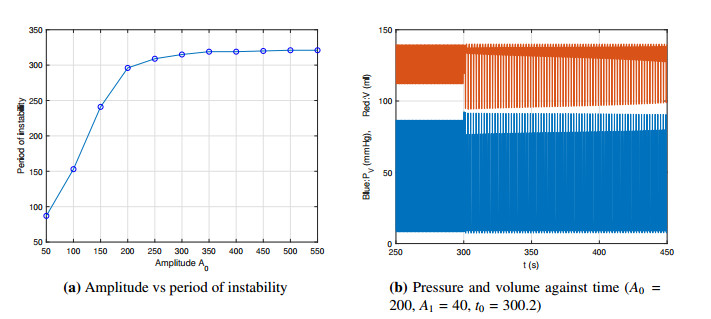
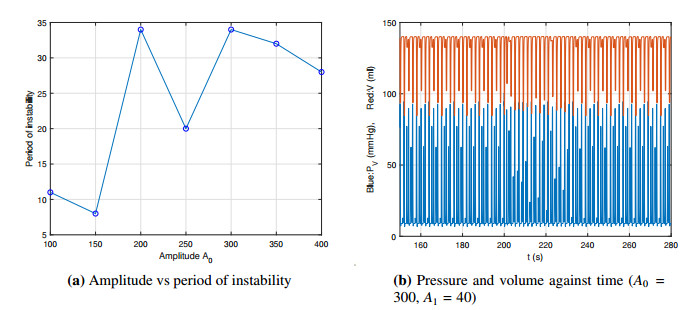
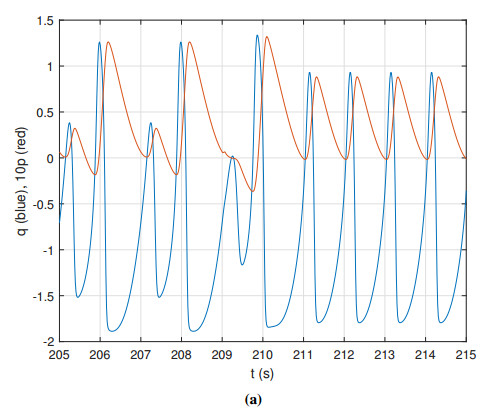
Figures(14) / Tables(2)

Nicholas Pearce, Eun-jin Kim. Modelling the cardiac response to a mechanical stimulation using a low-order model of the heart[J]. Mathematical Biosciences and Engineering, 2021, 18(4): 4871-4893. doi: 10.3934/mbe.2021248







 DownLoad:
DownLoad: