Citation: Alessia Andò, Dimitri Breda, Giulia Gava. How fast is the linear chain trick? A rigorous analysis in the context of behavioral epidemiology[J]. Mathematical Biosciences and Engineering, 2020, 17(5): 5059-5084. doi: 10.3934/mbe.2020273

| [1] | F. Curtain, H. Zwart, An Introduction to Infinite-Dimensional Linear Systems Theory, Texts in Applied Mathematics 21, Springer-Verlag, New York, 1995. |
| [2] | O. Diekmann, M. Gyllenberg, J. A. J. Metz, Finite dimensional state representation of linear and nonlinear delay systems, J. Dynam. Differ. Equations, 30 (2018), 1439-1467. |
| [3] | A. K. Erlang, Solution of some problems in the theory of probabilities of significance in automatic telephone exchanges, Post Office Elec. Eng., (1917), 189-197. |
| [4] | N. MacDonald, Time Lags in Biological Models, Lecture Notes in Biomathematics 27, Springer Verlag, Berlin, 1978. |
| [5] | N. MacDonald, Biological Delay Systems: Linear Stability Theory, Cambridge Studies in Mathematical Biology 8, Cambridge Univeristy Press, Cambridge, 1989. |
| [6] | D. Fargue, Reductibilitè des systèmes héréditaires à des systèmes dynamiques, C.R. Acad. Sci. Paris Sér. A-B, 277 (1973), 471-473. |
| [7] | D. Fargue, Reductibilitè des systèmes héréditaires, Int. J. Nonlin. Mech., 9 (1974), 331-338. |
| [8] | T. Vogel, Théorie Des Systèmes Evolutifs, Gautier Villars, Paris, 1965. |
| [9] | D. Breda, O. Diekmann, M. Gyllenberg, F. Scarabel, R. Vermiglio, Pseudospectral discretization of nonlinear delay equations: New prospects for numerical bifurcation analysis, SIAM J. Appl. Dyn. Sys., 15 (2016), 1-23. |
| [10] | S. Busenberg, C. Travis, On the use of reducible-functional differential equations in biological models, J. Math. Anal. Appl., 89 (1982), 46-66. |
| [11] | K. L. Cooke, Z. Grossman, Discrete delay, distributed delays and stability switches, J. Math. Anal. Appl., 86 (1982), 592-627. |
| [12] | E. Beretta, D. Breda, Discrete or distributed delay? Effects on stability of population growth, Math. Biosci. Eng., 13 (2016), 19-41. |
| [13] | C. Barril, A. Calsina, J. Ripoll, A practical approach to R0 in continuous-time ecological models, Math. Meth. Appl. Sci., 41 (2018), 8432-8445. |
| [14] | A. Lloyd, Destabilization of epidemic models with the inclusion of realistic distributions of infectious periods, Proc. Roy. Soc. Lond. B, 268 (2001), 985-993. |
| [15] | A. Lloyd, Realistic distributions of infectious periods in epidemic models: Changing patterns of persistence and dynamics, Theor. Popul. Biol., 60 (2001), 59-71. |
| [16] | C. Bauch, A. d'Onofrio, P. Manfredi, Behavioral epidemiology of infectious diseases: An overview, in Modeling the Interplay Between Human Behavior and the Spread of Infectious Diseases (eds. P. Manfredi and A. d'Onofrio), Springer-Verlag, New York, (2013), 1-19. |
| [17] | P. Manfredi, A. d'Onofrio, Modeling the Interplay Between Human Behavior and the Spread of Infectious Diseases, Springer-Verlag, New York, 2013. |
| [18] | Z. Wang, T. C. Bauch, S. Bhattacharyya, A. d'Onofrio, P. Manfredi, M. Percg, et al., Statistical physics of vaccination, Phys. Rep., 664 (2016), 1-113. |
| [19] | B. Buonomo, G. Carbone, A. d'Onofrio, Effect of seasonality on the dynamics of an imitationbased vaccination model with public health intervention, Math. Biosci., 15 (2018), 299-321. |
| [20] | B. Buonomo, A. d'Onofrio, D. Lacitignola, Global stability of an sir epidemic model with information dependent vaccination, Math. Biosci., 216 (2008), 9-16. |
| [21] | B. Buonomo, A. d'Onofrio, D. Lacitignola, The geometric approach to global stability in behavioral epidemiology, in Modeling the Interplay Between Human Behavior and the Spread of Infectious Diseases (eds. P. Manfredi and A. d'Onofrio), Springer-Verlag, New York, (2013), 289-308. |
| [22] | A. d'Onofrio, P. Manfredi, Information-related changes in contact patterns may trigger oscillations in the endemic prevalence of infectious diseases, J. Theoret. Biol., 256 (2009), 473-478. |
| [23] | A. d'Onofrio, P. Manfredi, Vaccine demand driven by vaccine side effects: Dynamic implications for sir diseases, J. Theoret. Biol., 264 (2010), 237-252. |
| [24] | A. d'Onofrio, P. Manfredi, P. Poletti, The impact of vaccine side effects on the natural history of immunization programmes: an imitation-game approach, J. Theoret. Biol., 273 (2011), 63-71. |
| [25] | A. d'Onofrio, P. Manfredi, E. Salinelli, Vaccinating behaviour and the dynamics of vaccine preventabe infections, in Modeling the Interplay Between Human Behavior and the Spread of Infectious Diseases (eds. P. Manfredi and A. d'Onofrio), Springer-Verlag, New York, (2013), 267-287. |
| [26] | S. Funk, M. Salathé, V. A. Jansen, Modelling the influence of human behaviour on the spread of infectious diseases: A review, J. R. Soc. Interface, 7 (2010), 1247-1256. |
| [27] | A. d'Onofrio, P. Manfredi, E. Salinelli, Vaccinating behaviour, information, and the dynamics of SIR vaccine preventable diseases, Theor. Popul. Biol., 71 (2007), 301-317. |
| [28] | A. d'Onofrio, Mixed pulse vaccination strategy in epidemic model with realistically distributed infectious and latent times, Appl. Math. Comput, 151 (2004), 181-187. |
| [29] | A. Calsina, J. Ripoll, Hopf bifurcation in a structured population model for the sexual phase of monogonont rotifers, J. Math. Biol., 45 (2002), 22-36. |
| [30] | D. Breda, O. Diekmann, S. Maset, R. Vermiglio, A numerical approach for investigating the stability of equilibria for structured population models, J. Biol. Dyn., 7 (2013), 4-20. |
| [31] | D. Breda, S. Maset, R. Vermiglio, Pseudospectral differencing methods for characteristic roots of delay differential equations, SIAM J. Sci. Comput., 27 (2005), 482-495. |
| [32] | D. Breda, S. Maset, R. Vermiglio, Stability of Linear Delay Differential Equations-A Numerical Approach with MATLAB, Springer, New York, 2015. |
| [33] | N. Olgac, R. Sipahi, Kernel and offspring concepts for the stability robustness of multiple time delayed systems (MTDS), J. Dyn. Syst. T. ASME, 129 (2006), 245-251. |
| [34] | O. Diekmann, P. Getto, M. Gyllenberg, Stability and bifurcation analysis of Volterra functional equations in the light of suns and stars, SIAM J. Math. Anal., 39 (2008), 1023-1069. |
| [35] | D. Breda, P. Getto, J. Sánchez Sanz, R. Vermiglio, Computing the eigenvalues of realistic Daphnia models by pseudospectral methods, SIAM J. Sci. Comput., 37 (2015), 2607-2629. |
| [36] | H. A. Priestley, Introduction to Complex Analysis, Oxford University Press, New York, 1990. |
| [37] | L. Fanti, P. Manfredi, The Solow's model with endogenous population: A neoclassical growth cycle model, J. Econ. Dev., 28 (2003), 103-115. |
| [38] | P. Manfredi, L. Fanti, Cycles in dynamic economic modelling, Econ. Model., 21 (2004), 573-594. |
| [39] | D. Breda, D. Liessi, Approximation of eigenvalues of evolution operators for linear renewal equations, SIAM J. Numer. Anal., 56 (2018), 1456-1481. |
| [40] | D. Breda, S. Maset, R. Vermiglio, Approximation of eigenvalues of evolution operators for linear retarded functional differential equations, SIAM J. Numer. Anal., 50 (2012), 1456-1483. |
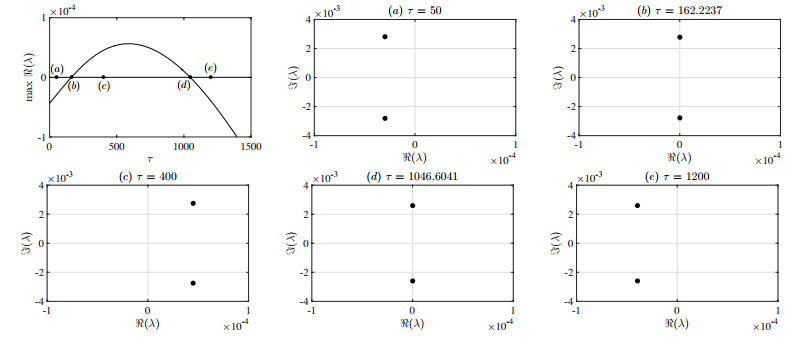
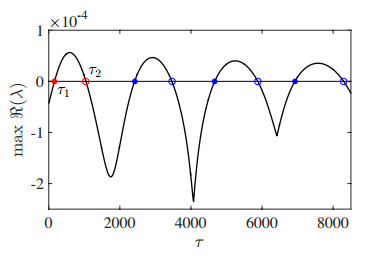
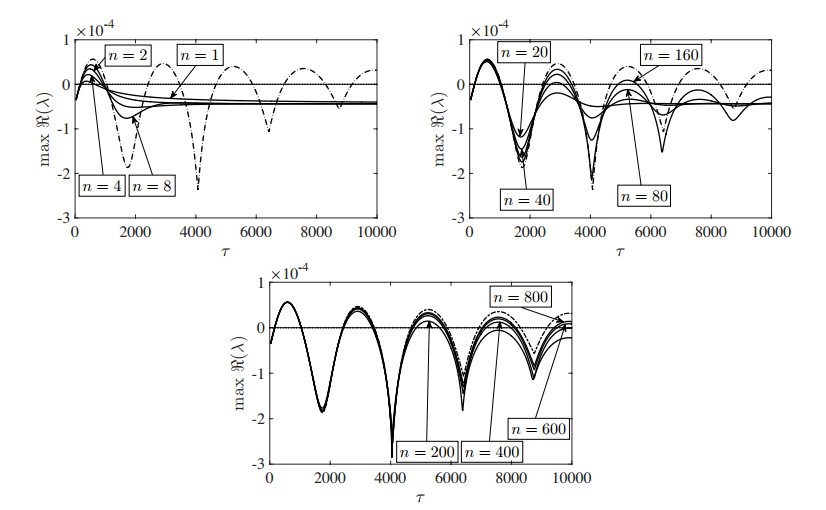

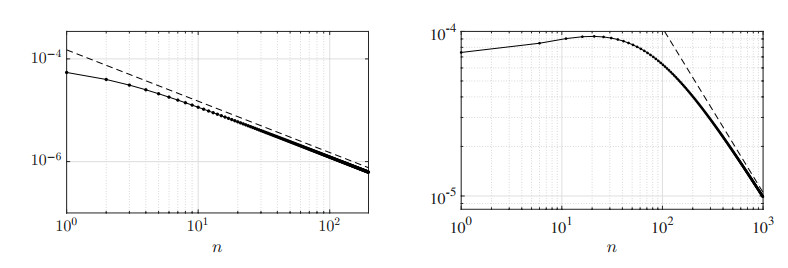
Figures(13) / Tables(3)

Alessia Andò, Dimitri Breda, Giulia Gava. How fast is the linear chain trick? A rigorous analysis in the context of behavioral epidemiology[J]. Mathematical Biosciences and Engineering, 2020, 17(5): 5059-5084. doi: 10.3934/mbe.2020273







 DownLoad:
DownLoad: