Citation: Anshuman Swain, William F Fagan. A mathematical model of the Warburg Effect: Effects of cell size, shape and substrate availability on growth and metabolism in bacteria[J]. Mathematical Biosciences and Engineering, 2019, 16(1): 168-186. doi: 10.3934/mbe.2019009

| [1] | M. Basan, S. Hui, H. Okano, Z. Zhang, Y. Shen, J. R. Williamson and T. Hwa, Overflow metabolism in Escherichia coli results from e_cient proteome allocation, Nature, 528 (2015) 99–104. |
| [2] | M. Bekker, S. de Vries, A. Ter Beek, K. J. Hellingwerf and M. J. T. de Mattos, Respiration of Escherichia coli Can Be Fully Uncoupled via the Nonelectrogenic Terminal Cytochrome bd-II Oxidase, J. Bacteriol., 191 (2009), 5510–5517. |
| [3] | Q. K. Beg, A. Vazquez, J. Ernst, M. A. de Menezes, Z. Bar-Joseph, A. L. Barabasi, and Z. N. Oltvai Intracellular crowding defines the mode and sequence of substrate uptake by Escherichia coli and constrains its metabolic activity, Proc. Natl. Acad. Sci. U S A, 104 (2007), 12663–12668. |
| [4] | K. J. Begg and W. D. Donachie, Cell shape and division in Escherichia coli: experiments with shape and division mutants, J. Bacteriol., 163 (1985), 615–622. |
| [5] | E. M. Corteselli, J. C. Burtis, A. K. Heinz and J. B. Yavitt, Leaf Litter Fuels Methanogenesis Throughout Decomposition in a Forested Peatland, Ecosystems, 20 (2017), 1217–1232. |
| [6] | J. P. DeLong, J. G. Okie, M. E. Moses, R. M. Sibly and J. H. Brown, Shifts in metabolic scaling, production, and e_ciency across major evolutionary transitions of life, Proc. Natl. Acad. Sci. U.S.A, 107 (2010), 12941–12945. |
| [7] | R. H. de Deken, The Crabtree E_ect: A Regulatory System in Yeast, Microbiology, 44 (1966), 149–156. |
| [8] | C. E. Deutch and G. S. Perera, Myceloid cell formation in Arthrobacter globiformis during osmotic stress, J. Appl. Bacteriol., 72 (1992), 493–499. |
| [9] | A. Esteve-Nunez, M. Rothermich, M. Sharma and D. Lovley, Growth of Geobacter sulfurreducens under nutrient-limiting conditions in continuous culture, Environ. Microbiol., 7 (2005), 641–648. |
| [10] | I. Famili, J. Forster, J. Nielson and B. O. Palsson, Saccharomyces cerevisiae phenotypes can be predicted by using constraint-based analysis of a genome-scale reconstructed metabolic network, Proc. Natl. Acad. Sci. U.S.A, 100 (2003), 13134–13139. |
| [11] | A. Flamholz, E. Noor, A. Bar-Even,W. Liebermeister and R. Milo, Tradeo_s in glycolytic strategy, Proc. Natl. Acad. Sci. U.S.A, 110 (2013), 10039–10044. |
| [12] | A. Frenkel and W. Hirsch, Spontaneous development of L forms of streptococci requiring secretions of other bacteria or sulphydryl compounds for normal growth, Nature, 191 (1961), 728–730. |
| [13] | T. Frick and S. Schuster, An example of the prisoner's dilemma in biochemistry, Naturwis-senschaften, 90 (2003), 327–331. |
| [14] | J. J. Germida and L. E. Casida Jr, Myceloid growth of Arthrobacter globiformis and other Arthrobacter species, J. Bacteriol., 144 (1980), 1152–1158. |
| [15] | G. Jayachandran, H. Gorisch and L. Adrian, Studies on hydrogenase activity and chlorobenzene respiration in Dehalococcoides sp. strain CBDB1, Arch. Microbiol., 182 (2004), 498–504. |
| [16] | I. Kareva, Prisoner's dilemma in cancer metabolism, PLoS One, 6 (2011), e28576. |
| [17] | C. P. Kempes, P. M. van Bodegom, D. Wolpert, E. Libby, J. Amend, and T. Hoehler, Drivers of Bacterial Maintenance and Minimal Energy Requirements, Front. Microbiol., 8 (2017), 31. |
| [18] | R. Lange and R. Hengge-Aronis, Growth phase-regulated expression of bolA and morphology of stationary-phase Escherichia coli cells are controlled by the novel sigma factor sigma S, J. Bacteriol., 173 (1991), 4474–4481. |
| [19] | R. E. Lenski and M. Travisano, Dynamics of adaptation and diversification: a 10,000-generation experiment with bacterial populations, Proc. Natl. Acad. Sci. U.S.A, 91 (1994), 6808–6814. |
| [20] | S. Y. Lunt and M. G. Vander Heiden, Aerobic Glycolysis: Meeting the Metabolic Requirements of Cell Proliferation, Annu. Rev. Cell Dev. Biol., 27 (2011), 441–464. |
| [21] | M. J. McInerney and M. P. Bryant, Anaerobic Degradation of Lactate by Syntrophic Associations of Methanosarcina barkeri and Desulfovibrio Species and Effect of H2 on Acetate Degradation, Appl. Environ. Microbiol., 41 (1981), 346–354. |
| [22] | D. Molenaar, R. van Berlo, D. de Ridder and B. Teusink, Shifts in growth strategies reflect tradeoffs in cellular economics, Mol. Syst. Biol., 5 (2009), 323. |
| [23] | P. M¨oller, X. Liu, S. Schuster and D. Boley, Linear programming model can explain respiration of fermentation products, PLoS ONE 13 (2018), e0191803. |
| [24] | N. Nanninga, Growth and form in microorganisms: morphogenesis of Escherichia coli, Can. J. Microbiol., 34 (1988), 381–389. |
| [25] | E. A. Newsholme, B. Crabtree and M. S. Ardawi The role of high rates of glycolysis and glutamine utilization in rapidly dividing cells, Biosc. Rep., 5 (1985), 393–400. |
| [26] | T. Pfei_er, S. Schuster and S. Bonhoe_er, Cooperation and competition in the evolution of ATPproducing pathways, Science, 292 (2001), 504–507. |
| [27] | L. Pine and C. J. Boone, Comparative cell wall analyses of morphological forms within the genus Actinomyces, J. Bacteriol., 94 (1967), 875–883. |
| [28] | N. D. Price, J. L. Reed and B. O. Palsson, Genome-scale models of microbial cells: evaluating the consequences of constraints, Nat. Rev. Microbiol., 2 (2004), 886–897. |
| [29] | C. Risso, J. Sun, K. Zhuang, R. Mahadevan, R. DeBoy, W. Ismail, S. Shrivastava, H. Huot, S. Kothari, S. Daugherty, O. Bui, C. H. Schilling, D. R. Lovley and B. A. Methe, Genome-scale comparison and constraint-based metabolic reconstruction of the facultative anaerobic Fe(III)-reducer Rhodoferax ferrireducens, BMC Genomics, 10 (2009), 447. |
| [30] | D. H. Rothman, G. P. Fournier, K. L. French, E. J. Alm, E. A. Boyle, C. Cao and R. E. Summons, Methanogenic burst in the end-Permian carbon cycle, Proc. Natl. Acad. Sci. U.S.A, 111 (2014), 5462–5467. |
| [31] | K. L. Ruoff, Nutritionally variant streptococci, Clin. Microbiol. Rev., 4 (1991), 184–190. |
| [32] | E. Ruppin, J. A. Papin, L. F. de Figueiredo and S. Schuster, Metabolic reconstruction, constraintbased analysis and game theory to probe genome-scale metabolic networks, Curr. Opin. Biotech., 21 (2010), 502–510. |
| [33] | M. Schaechter, O. Maaloe and N. O. Kjeldgaard, Dependency on medium and temperature of cell size and chemical composition during balanced growth of Salmonella typhimurium, J. Gen. Microbiol., 19 (1958), 592–606. |
| [34] | S. Schuster, D. Boley, P. Moller, H. Stark and C. Kaleta, Mathematical models for explaining the Warburg e_ect: a review focussed on ATP and biomass production, Biochem. Soc. Trans., 43 (2015), 1187–1194. |
| [35] | T. E. Shehata and A. G Marr, E_ect of Nutrient Concentration on the Growth of Escherichia coli, J. Bacteriol., 107 (1971), 210–216. |
| [36] | H. Shim and S. T. Yang, Biodegradation of benzene, toluene, ethylbenzene, and o-xylene by a coculture of Pseudomonas putida and Pseudomonas fluorescens immobilized in a fibrous-bed bioreactor, J. Biotechnol., 67 (1999), 99–112. |
| [37] | T. Shlomi, T. Benyamini, E. Gottlieb, R. Sharan and E. Ruppin, Genome-scale metabolic modeling elucidates the role of proliferative adaptation in causing the Warburg effect, PLoS Comput. Biol., 7 (2011), e1002018. |
| [38] | R. E. Steinberger, A. R. Allen, H. G. Hansma and P. A. Holden, Elongation correlates with nutrient deprivation in Pseudomonas aeruginosa unsaturated biofilms, Microb. Ecol., 43 (2002), 416–423. |
| [39] | A. Swain and S. Chatterjee, A new formulation for determination of the competition coeffcient in multispecies interaction for Lotka-Volterra type competition models, Curr. Sci., 112 (2017), 1920–1926. |
| [40] | C. P. Tseng, J. Albrecht and R. P. Gunsalus, Effect of microaerophilic cell growth conditions on expression of the aerobic (cyoABCDE and cydAB) and anaerobic (narGHJI, frdABCD, and dmsABC) respiratory pathway genes in Escherichia coli, J. Bacteriol., 178 (1996), 1094–1098. |
| [41] | M. G. Vander Heiden, L. C. Cantley and C. B. Thompson, Understanding the Warburg effect: the metabolic requirements of cell proliferation, Science, 324 (2009), 1029–1033. |
| [42] | A. Vazquez, J. Liu, Y. Zhou and Z. N. Oltvai Catabolic effciency of aerobic glycolysis: The Warburg effect revisited, BMC Syst. Biol., 4 (2010), 58. |
| [43] | G. N. Vemuri, E. Altman, D. P. Sangurdekar, A. B. Khodursky and M. A. Eiteman, Overflow metabolism in Escherichia coli during steady-state growth: transcriptional regulation and effect of the redox ratio, Appl. Environ. Microbiol., 72 (2006), 3653–3661. |
| [44] | D. Voet and J. G. Voet, Biochemistry, 3rd edition, Wiley and Sons, Hoboken, 2004. |
| [45] | O. Warburg, Origin of cancer cells, Science, 123 (1956), 309–314. |
| [46] | M. Webb, The influence of magnesium on cell division. I. The growth of Clostridium welchii in complex media deficient in magnesium, J. Gen. Microbiol., 2 (1948), 275–287. |
| [47] | M. Webb, E_ects of magnesium on cellular division in bacteria, Science, 118 (1953), 607–611. |
| [48] | K. D. Young, The selective value of bacterial shape, Microbiol. Mol. Biol. Rev., 70 (2006), 660–703. |
| [49] | K. Zhuang, G. N. Vemuri and R. Mahadevan, Economics of membrane occupancy and respirofermentation, Mol. Syst. Biol., 7 (2011), 500. |
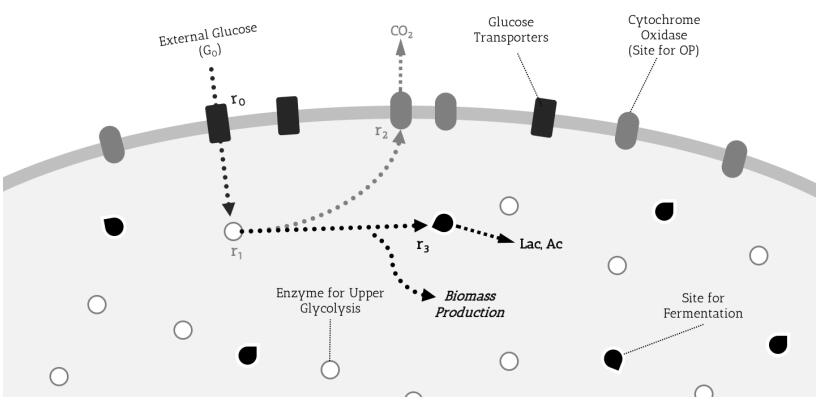
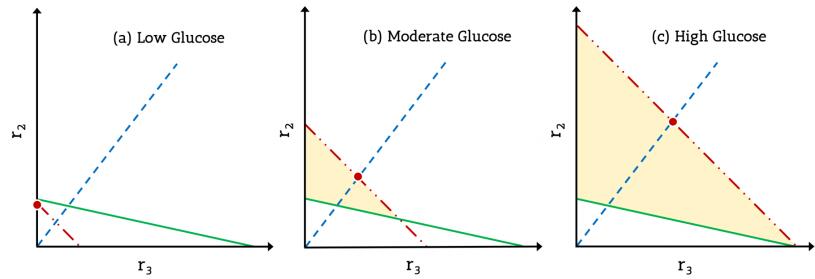
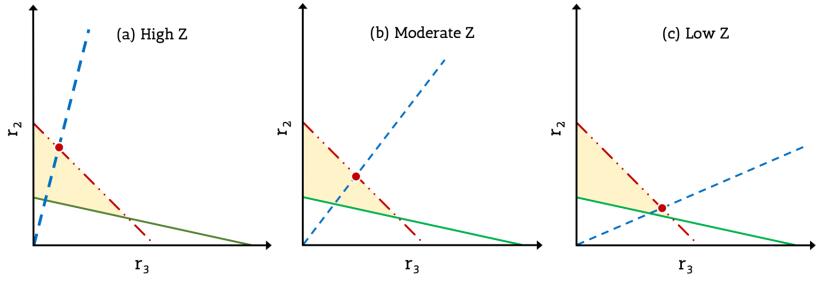
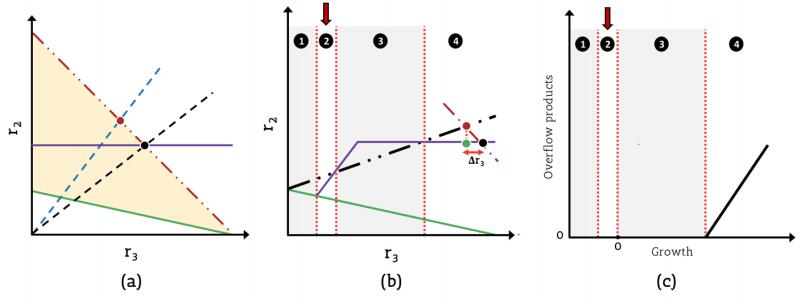
Figures(5) / Tables(1)

Anshuman Swain, William F Fagan. A mathematical model of the Warburg Effect: Effects of cell size, shape and substrate availability on growth and metabolism in bacteria[J]. Mathematical Biosciences and Engineering, 2019, 16(1): 168-186. doi: 10.3934/mbe.2019009







 DownLoad:
DownLoad: