Citation: Da Song, Meng Fan, Ming Chen, Hao Wang. Dynamics of a periodic stoichiometric model with application in predicting and controlling algal bloom in Bohai Sea off China[J]. Mathematical Biosciences and Engineering, 2019, 16(1): 119-138. doi: 10.3934/mbe.2019006

| [1] | G. Ahlgren, Temperature functions in biology and their application to algal growth constants,. Oikos, 49 (1987), 177–190. |
| [2] | J.H. Landsberg, The effects of harmful algal blooms on aquatic organisms, Rev. Fisher. Sci., 10 (2002), 113–390. |
| [3] | X. Li and H.Wang, A stoichiometrically derived algal growth model and its global analysis, Math. Biosci. Eng., 7 (2010), 825–836. |
| [4] | Y. Qi, Z. Zhang, Y. Hong, S. Lu, C. Zhu, and Y. Li, Occurrence of red tides on the coasts of China, In T.J. Smayda, and Y. Shimizu (Eds.) Toxic Marine Phytoplankton, Elsevier, Amsterdam, (1993), 43–46. |
| [5] | S. Tan, G. Shi, J. Shi, H. Gao, and X. Yao, Correlation of Asian dust with chlorophyll and primary productivity in the coastal seas of China during the period from 1998 to 2008, J. Geophys. Res., 116 (2011), doi:10.1029/2010JG001456. |
| [6] | C.K. Tseng, M.J. Zhou, and J.Z. Zou, Toxic phytoplankton studies in China, In T. J. Smayda, and Y. Shimizu (Eds.) Toxic Marine Phytoplankton. Elsevier, Amsterdam, (1993), 347–452. |
| [7] | T. Yan, M.J. Zhou, and J.Z. Zou, A national report on harmful algal blooms in China, In: Taylor “Max”, F. J. R. & V. L. Trainer (eds.). Harmful algal blooms in the PICES region of the North Pacific. PICES Scientific Report No. 23. North Pacific Marine Science Organization, Sidney, BC, Canada, (2002), 119–128. |
| [8] | R.E. Gaines and J.L. Mawhin, Coincidence Degree and Nonlinear Differential Equations, Springer-Verlag, Berlin, 1977. |
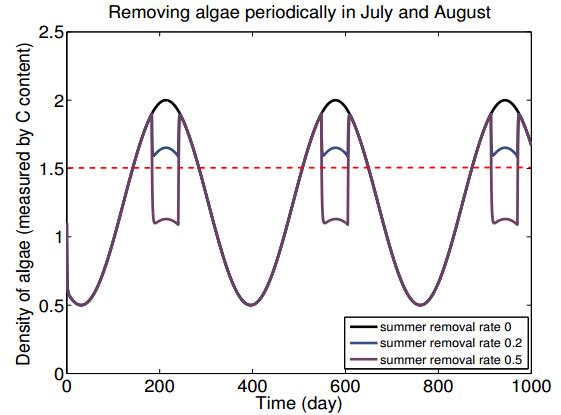
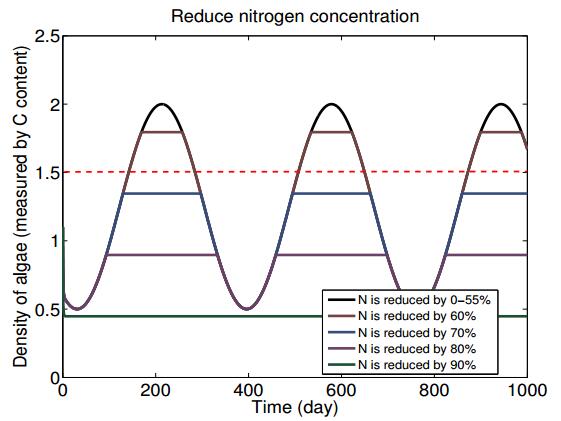
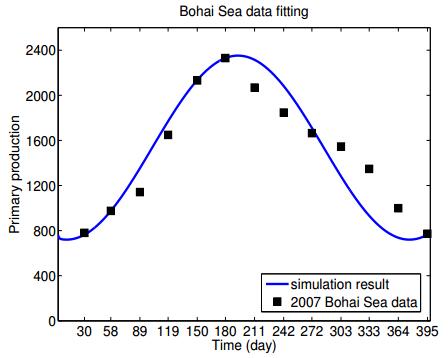
Figures(7) / Tables(2)

Da Song, Meng Fan, Ming Chen, Hao Wang. Dynamics of a periodic stoichiometric model with application in predicting and controlling algal bloom in Bohai Sea off China[J]. Mathematical Biosciences and Engineering, 2019, 16(1): 119-138. doi: 10.3934/mbe.2019006







 DownLoad:
DownLoad: