Citation: Neha Mittal, Anand kumar Srivastava. Cd2++Cr6+causes toxic effects on chromosomal development of microspore in Carthamus tinctorius[J]. AIMS Genetics, 2019, 6(1): 1-10. doi: 10.3934/genet.2019.1.1

| [1] |
Modliszewski JL, Wang H, Albright AR, et al. (2018) Elevated temperature increases meiotic crossover frequency via the interfering (Type I) pathway in Arabidopsis thaliana. PLoS Genet 14: e1007384. doi: 10.1371/journal.pgen.1007384

|
| [2] |
Somashekhar RK (1987) Meiotic abnormalities induced by dye industry wastewater in Chlorophytum amaniense Engler. Cytologia 52: 647–652. doi: 10.1508/cytologia.52.647
|
| [3] | Ramel C (1972) Genetic effects. In: Friberg L, Vostal D, Mercury in the Environment, Clevel and Ohio: CRC Press, 169–181. |
| [4] | Sharma DK (2001) Analysis of Genetic Variability and Inheritance for Resistance to Organic Mercurial Toxicity in Certain Crops. Ph.D. thesis, CCS University, Meerut, India. |
| [5] | Tomar P (2004) Genetic Analyses for Tolerance to Pb2+ and Hg2+ in Wheat. Ph.D. thesis, CCS University, Meerut, India. |
| [6] | Madaan N (2002) Analysis of genetic variability and inheritance for some heavy metal tolerance in oil crop Carthamus tintorius L. Ph.D. thesis. Meerut University, Meerut, India. |
| [7] |
Mittal N, Srivastava (2014) Cadmium- and Chromium-induced aberrations in the reproductive biology of hordeum vulgare. Cytologia 79: 207–214. doi: 10.1508/cytologia.79.207
|
| [8] | Choudhary S, Mohammad Yunus, Khalil Ansari, et al. (2012) Cytotoxic action of lead nitrate on cytomorphology of Trigonella foenum-graceum L. Turk J Biol 36: 267–273. |
| [9] |
Morais DL, Marin-Morales MA (2009) Allium cepa test in environmental monitoring: A review on its application. Mutat Res 682: 71–81. doi: 10.1016/j.mrrev.2009.06.002
|
| [10] | Singh P (2015) Toxic effect of chromium on genotoxicity and cytotoxicity by use of Allium cepa L. Int J Res Eng App Sci 5: 1–10. |
| [11] | Sax K (1940) An analysis of X-ray induced chromosomal in Tradescantia. Genetics 25: 41–68. |
| [12] |
Shaikh MAQ, Godward MBE (1972) The mitotic consequences of radiation induced chromosome breaks in Lathyrus sativus and Vicia ervila. Cytologia 37: 497–505. doi: 10.1508/cytologia.37.497
|
| [13] |
Conger AD, Stevenson HO (1969) Correlation of seedling height and chromosomal damage in irradiated barely seeds. Radiat Bot 9: 1–14. doi: 10.1016/S0033-7560(69)80015-3
|
| [14] |
Mumthas S, Chidambaram ALA, Sundaramoorthy P, et al. (2010) Effect of arsenic and manganese on root growth and cell division in root tip cells of green gram (Vigna radiata L.). Emir J Food Agric 22: 285–297. doi: 10.9755/ejfa.v22i4.4876
|
| [15] | Kumar G, Tripathi R (2008) Lead-induced cytotoxicity and mutagenicity in grass pea. Turk J Biol 32: 73–78. |
| [16] | Rai PK, Kumar G (2010) The genotoxic potential of two heavy metals in inbred lines of maize (Zea mays L). Turk J Bot 34: 39–46. |
| [17] |
Gaulden ME (1987) Hypothesis some mutagens directly alter specific chromosomal proteins (DNA topoisomerase II and peripheral proteins) to produce chromosome stickiness, which causes chromosome aberrations. Mutagenesis 2: 357–365. doi: 10.1093/mutage/2.5.357
|
| [18] | Kumar G, Rai P (2007) Genotoxic potential of mercury and cadmium in soybean. Turk J Biol 31: 187–195. |
| [19] |
Patnaik S, Saran BL, Patnik SM (1984) Effect of Zarda (processed tobacco leaf) extract on the chromosome of Allium cepa. Cytologia 49: 807–814. doi: 10.1508/cytologia.49.807
|
| [20] | Rumana Aslam, Ansari MYK, Sana Choudhary, et al. (2014) Genotoxic effects of heavy metal cadmium on growth, biochemical, cyto-physiological parameters and detection of DNA polymorphism by RAPD in Capsicum annuumL.-An important spice crop of India. Saudi J Bio Sci 21: 465–472. |
| [21] | Auerbach C (1976) Mutation research problems. Results and perspective. Chapman and Hall London. |
| [22] | Kumar G, Srivastava S (2006) Buffering action of essential metals against heavy metal cytotoxicity in Helianthus aanus. Nucleus 49: 45–51. |
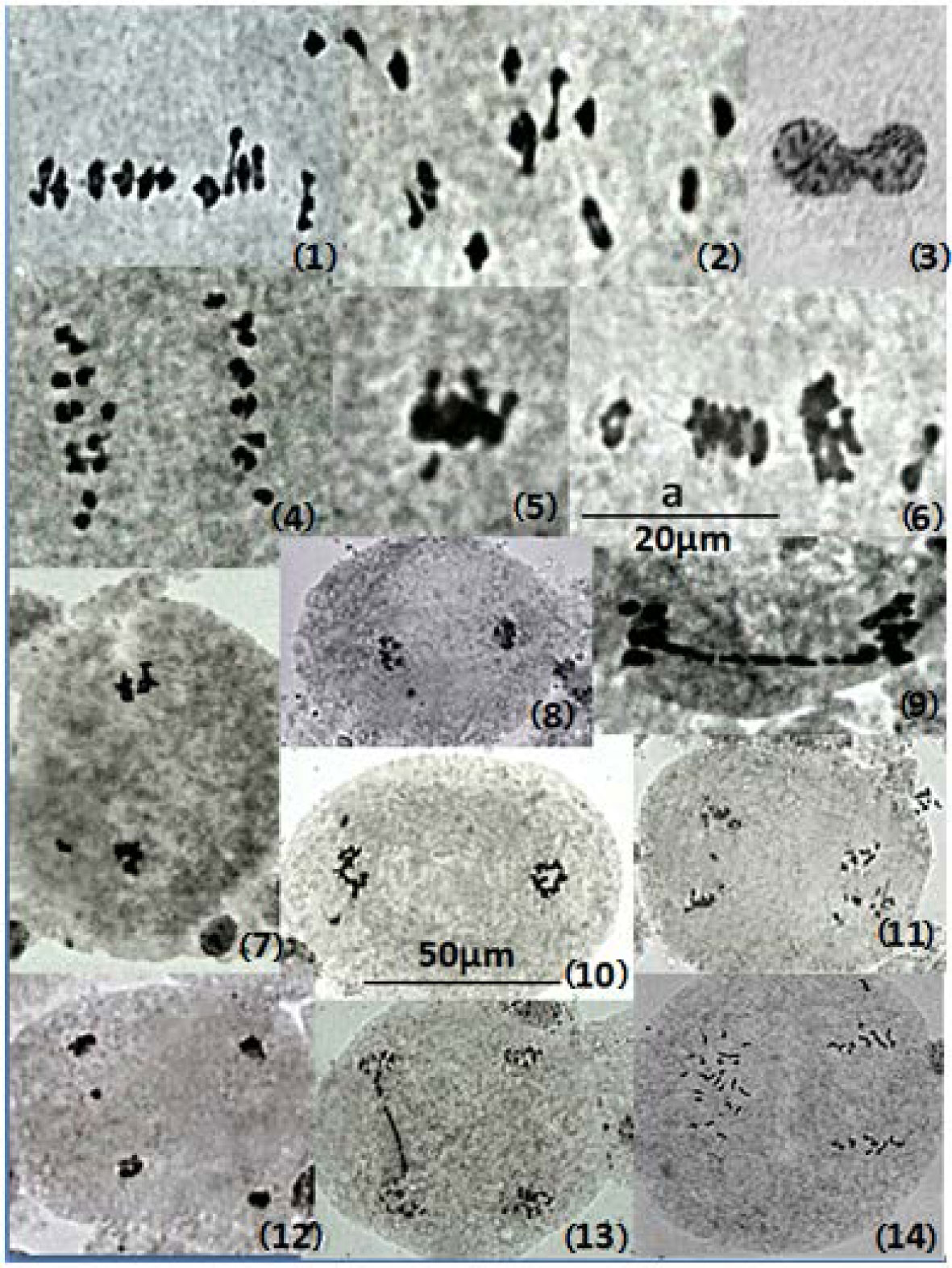
Figures(1) / Tables(4)

Neha Mittal, Anand kumar Srivastava. Cd2++Cr6+causes toxic effects on chromosomal development of microspore in Carthamus tinctorius[J]. AIMS Genetics, 2019, 6(1): 1-10. doi: 10.3934/genet.2019.1.1







 DownLoad:
DownLoad: