Digital holographic microscopy provides the ability to observe throughout a large volume without refocusing. This capability enables simultaneous observations of large numbers of microorganisms swimming in an essentially unconstrained fashion. However, computational tools for tracking large 4D datasets remain lacking. In this paper, we examine the errors introduced by tracking bacterial motion as 2D projections vs. 3D volumes under different circumstances: bacteria free in liquid media and bacteria near a glass surface. We find that while XYZ speeds are generally equal to or larger than XY speeds, they are still within empirical uncertainties. Additionally, when studying dynamic surface behavior, the Z coordinate cannot be neglected.
Citation: Jacqueline Acres, Jay Nadeau. 2D vs 3D tracking in bacterial motility analysis[J]. AIMS Biophysics, 2021, 8(4): 385-399. doi: 10.3934/biophy.2021030
Digital holographic microscopy provides the ability to observe throughout a large volume without refocusing. This capability enables simultaneous observations of large numbers of microorganisms swimming in an essentially unconstrained fashion. However, computational tools for tracking large 4D datasets remain lacking. In this paper, we examine the errors introduced by tracking bacterial motion as 2D projections vs. 3D volumes under different circumstances: bacteria free in liquid media and bacteria near a glass surface. We find that while XYZ speeds are generally equal to or larger than XY speeds, they are still within empirical uncertainties. Additionally, when studying dynamic surface behavior, the Z coordinate cannot be neglected.

| [1] |
Armitage JP (1997) Three hundred years of bacterial motility. Found Mod Biochem 3: 107-171. doi: 10.1016/S1874-5660(97)80007-X

|
| [2] |
Belas R, Zhulin IB, Yang Z (2008) Bacterial signaling and motility: sure bets. J Bacteriol 190: 1849-1856. doi: 10.1128/JB.01943-07
|
| [3] |
Holscher T, Bartels B, Lin YC, et al. (2015) Motility, chemotaxis and aerotaxis contribute to competitiveness during bacterial pellicle biofilm development. J Mol Biol 427: 3695-3708. doi: 10.1016/j.jmb.2015.06.014
|
| [4] |
Mitchell JG, Kogure K (2006) Bacterial motility: links to the environment and a driving force for microbial physics. FEMS Microbiol Ecol 55: 3-16. doi: 10.1111/j.1574-6941.2005.00003.x
|
| [5] |
Velho Rodrigues MF, Lisicki M, Lauga E (2021) The bank of swimming organisms at the micron scale (BOSO-Micro). Plos One 16: e0252291. doi: 10.1371/journal.pone.0252291
|
| [6] |
Vaituzis Z, Doetsch RN (1969) Motility tracks: technique for quantitative study of bacterial movement. Appl Microbiol 17: 584-588. doi: 10.1128/am.17.4.584-588.1969
|
| [7] |
Barnkob R, Rossi M (2020) General defocusing particle tracking: fundamentals and uncertainty assessment. Exp Fluids 61: 1-14. doi: 10.1007/s00348-020-2937-5
|
| [8] |
de Anda J, Lee EY, Lee CK, et al. (2017) High-speed “4D” computational microscopy of bacterial surface motility. Acs Nano 11: 9340-9351. doi: 10.1021/acsnano.7b04738
|
| [9] |
Lofroth M, Avci E (2018) Auto-focusing approach on multiple micro objects using the prewitt operator. Int J Intell Robot Appl 2: 413-424. doi: 10.1007/s41315-018-0070-x
|
| [10] |
Taute KM, Gude S, Tans SJ, et al. (2015) High-throughput 3D tracking of bacteria on a standard phase contrast microscope. Nat Commun 6: 8776. doi: 10.1038/ncomms9776
|
| [11] |
Qi M, Gong X, Wu B, et al. (2017) Landing dynamics of swimming bacteria on a polymeric surface: Effect of surface properties. Langmuir 33: 3525-3533. doi: 10.1021/acs.langmuir.7b00439
|
| [12] |
Yuan S, Qi M, Peng Q, et al. (2021) Adaptive behaviors of planktonic Pseudomonas aeruginosa in response to the surface-deposited dead siblings. Colloid Surface B 197: 111408. doi: 10.1016/j.colsurfb.2020.111408
|
| [13] |
Cheong FC, Wong CC, Gao YF, et al. (2015) Rapid, high-throughput tracking of bacterial motility in 3D via phase-contrast holographic video microscopy. Biophys J 108: 1248-1256. doi: 10.1016/j.bpj.2015.01.018
|
| [14] |
Farthing NE, Findlay RC, Jikeli JF, et al. (2017) Simultaneous two-color imaging in digital holographic microscopy. Opt Express 25: 28489-28500. doi: 10.1364/OE.25.028489
|
| [15] |
Flewellen JL, Zaid IM, Berry RM (2019) A multi-mode digital holographic microscope. Rev Sci Instrum 90: 023705. doi: 10.1063/1.5066556
|
| [16] | Hook AL, Flewellen JL, Dubern JF, et al. (2019) Simultaneous tracking of Pseudomonas aeruginosa motility in liquid and at the solid-liquid interface reveals differential roles for the flagellar stators. Msystems 4: e00390-19. |
| [17] |
Huang G, Tian WZ, Qi M, et al. (2018) Improving axial resolution for holographic tracking of colloids and bacteria over a wide depth of field by optimizing different factors. Opt Express 26: 9920-9930. doi: 10.1364/OE.26.009920
|
| [18] |
Rieu M, Vieille T, Radou G, et al. (2021) Parallel, linear, and subnanometric 3D tracking of microparticles with Stereo Darkfield Interferometry. Sci Adv 7: eabe3902. doi: 10.1126/sciadv.abe3902
|
| [19] |
Yamato K, Chiba H, Oku H (2020) High speed three dimensional tracking of swimming cell by synchronous modulation between tece camera and tag lens. IEEE Robot Autom Lett 5: 1907-1914. doi: 10.1109/LRA.2020.2969911
|
| [20] |
Bedrossian M, El-Kholy M, Neamati D, et al. (2018) A machine learning algorithm for identifying and tracking bacteria in three dimensions using Digital Holographic Microscopy. AIMS Biophys 5: 36-49. doi: 10.3934/biophy.2018.1.36
|
| [21] |
Cerda M, Navarro CA, Silva J, et al. (2018) A high-speed tracking algorithm for dense granular media. Comput Phys Commun 227: 8-16. doi: 10.1016/j.cpc.2018.02.010
|
| [22] |
Shao S, Mallery K, Kumar SS, et al. (2020) Machine learning holography for 3D particle field imaging. Opt Express 28: 2987-2999. doi: 10.1364/OE.379480
|
| [23] |
Wang GC, Huang G, Gong XJ, et al. (2020) Method for 3D tracking behaviors of interplaying bacteria individuals. Opt Express 28: 28060-28071. doi: 10.1364/OE.401032
|
| [24] |
Wang A, Garmann RF, Manoharan VN (2016) Tracking E-coli runs and tumbles with scattering solutions and digital holographic microscopy. Opt Express 24: 23719-23725. doi: 10.1364/OE.24.023719
|
| [25] |
Sheng J, Malkiel E, Katz J, et al. (2007) Digital holographic microscopy reveals prey-induced changes in swimming behavior of predatory dinoflagellates. P Natl Acad Sci USA 104: 17512-17517. doi: 10.1073/pnas.0704658104
|
| [26] |
Sheng J, Malkiel E, Katz J, et al. (2010) A dinoflagellate exploits toxins to immobilize prey prior to ingestion. P Natl Acad Sci USA 107: 2082-2087. doi: 10.1073/pnas.0912254107
|
| [27] |
Molaei M, Sheng J (2014) Imaging bacterial 3D motion using digital in-line holographic microscopy and correlation-based de-noising algorithm. Opt Express 22: 32119-32137. doi: 10.1364/OE.22.032119
|
| [28] |
Wang A, Garmann RF, Manoharan VN (2016) Tracking E. coli runs and tumbles with scattering solutions and digital holographic microscopy. Opt Express 24: 23719-23725. doi: 10.1364/OE.24.023719
|
| [29] |
Yu X, Hong J, Liu C, et al. (2014) Review of digital holographic microscopy for three-dimensional profiling and tracking. Opt Eng 53: 112306. doi: 10.1117/1.OE.53.11.112306
|
| [30] |
Stocker R (2011) Reverse and flick: Hybrid locomotion in bacteria. P Natl Acad Sci USA 108: 2635-2636. doi: 10.1073/pnas.1019199108
|
| [31] |
Bubendorfer S, Held S, Windel N, et al. (2012) Specificity of motor components in the dual flagellar system of Shewanella putrefaciens CN-32. Mol Microbiol 83: 335-350. doi: 10.1111/j.1365-2958.2011.07934.x
|
| [32] |
Butler SM, Camilli A (2004) Both chemotaxis and net motility greatly influence the infectivity of Vibrio cholerae. P Natl Acad Sci USA 101: 5018-5023. doi: 10.1073/pnas.0308052101
|
| [33] |
Butler SM, Camilli A (2005) Going against the grain: chemotaxis and infection in Vibrio cholerae. Nat Rev Microbiol 3: 611-620. doi: 10.1038/nrmicro1207
|
| [34] |
Paździor E, Pękala-Safińska A, Wasyl D (2019) Phenotypic diversity and potential virulence factors of the Shewanella putrefaciens group isolated from freshwater fish. J Vet Res 63: 321-332. doi: 10.2478/jvetres-2019-0046
|
| [35] |
Pękala A, Kozińska A, Paździor E, et al. (2015) Phenotypical and genotypical characterization of Shewanella putrefaciens strains isolated from diseased freshwater fish. J Fish Dis 38: 283-293. doi: 10.1111/jfd.12231
|
| [36] |
Vignier N, Barreau M, Olive C, et al. (2013) Human infection with Shewanella putrefaciens and S. algae: report of 16 cases in Martinique and review of the literature. Am J Trop Med Hyg 89: 151-156. doi: 10.4269/ajtmh.13-0055
|
| [37] |
Kühn J, Niraula B, Liewer K, et al. (2014) A Mach-Zender digital holographic microscope with sub-micrometer resolution for imaging and tracking of marine micro-organisms. Rev Sci Instrum 85: 123113. doi: 10.1063/1.4904449
|
| [38] |
Nadeau JL, Cho YB, Kühn J, et al. (2016) Improved tracking and resolution of bacteria in holographic microscopy using dye and fluorescent protein labeling. Front Chem 4: 17. doi: 10.3389/fchem.2016.00017
|
| [39] | Fregoso SF, Lima F, Wallace JK, et al. (2020) DHMx (Digital holographic microscope experience) software tool, Digital Holography and Three-Dimensional Imaging. Opt Soc Am HF2G.7. |
| [40] | Mann CJ, Kim MK (2006) Quantitative phase-contrast microscopy by angular spectrum digital holography, Three-Dimensional and Multidimensional Microscopy: Image Acquisition and Processing XIII. International Society for Optics and Photonics 6090: 60900B. |
| [41] |
Cohoe D, Hanczarek I, Wallace JK, et al. (2019) Multiwavelength imaging and unwrapping of protozoa in amplitude and phase using custom Fiji plug-ins. Frontiers Phys 7: 94. doi: 10.3389/fphy.2019.00094
|
| [42] |
Colomb T, Kühn J, Charrière F, et al. (2006) Total aberrations compensation in digital holographic microscopy with a reference conjugated hologram. Opt Express 14: 4300-4306. doi: 10.1364/OE.14.004300
|
| [43] |
Schindelin J, Arganda-Carreras I, Frise E, et al. (2012) Fiji: an open-source platform for biological-image analysis. Nat Methods 9: 676-682. doi: 10.1038/nmeth.2019
|
| [44] |
Taute KM, Gude S, Tans SJ, et al. (2015) High-throughput 3D tracking of bacteria on a standard phase contrast microscope. Nat Commun 6: 8776. doi: 10.1038/ncomms9776
|
| [45] |
Vater SM, Weiße S, Maleschlijski S, et al. (2014) Swimming behavior of Pseudomonas aeruginosa studied by holographic 3D tracking. PLoS One 9: e87765. doi: 10.1371/journal.pone.0087765
|
| [46] |
Heydt M, Divós P, Grunze M, et al. (2009) Analysis of holographic microscopy data to quantitatively investigate three-dimensional settlement dynamics of algal zoospores in the vicinity of surfaces. Eur Phys J E 30: 141-148. doi: 10.1140/epje/i2009-10459-9
|
| [47] |
Dixon L, Cheong FC, Grier DG (2011) Holographic deconvolution microscopy for high-resolution particle tracking. Opt Express 19: 16410-16417. doi: 10.1364/OE.19.016410
|
| [48] |
Latychevskaia T, Fink HW (2014) Holographic time-resolved particle tracking by means of three-dimensional volumetric deconvolution. Opt Express 22: 20994-21003. doi: 10.1364/OE.22.020994
|
| [49] |
Goto T, Nakata K, Baba K, et al. (2005) A fluid-dynamic interpretation of the asymmetric motion of singly flagellated bacteria swimming close to a boundary. Biophys J 89: 3771-3779. doi: 10.1529/biophysj.105.067553
|
| [50] |
Lauga E, Diluzio WR, Whitesides GM, et al. (2006) Swimming in circles: Motion of bacteria near solid boundaries. Biophys J 90: 400-412. doi: 10.1529/biophysj.105.069401
|
| [51] |
Das D, Lauga E (2019) Transition to bound states for bacteria swimming near surfaces. Phys Rev E 100: 043117. doi: 10.1103/PhysRevE.100.043117
|
| [52] |
Kudo S, Imai N, Nishitoba M, et al. (2005) Asymmetric swimming pattern of Vibrio alginolyticuscells with single polar flagella. FEMS Microbiol Lett 242: 221-225. doi: 10.1016/j.femsle.2004.11.007
|
| [53] |
Chen M, Zhao Z, Yang J, et al. (2017) Length-dependent flagellar growth of Vibrio alginolyticus revealed by real time fluorescent imaging. ELife 6: e22140. doi: 10.7554/eLife.22140
|
| [54] |
Molaei M, Barry M, Stocker R, et al. (2014) Failed escape: Solid surfaces prevent tumbling of Escherichia coli. Phys Rev Lett 113: 068103. doi: 10.1103/PhysRevLett.113.068103
|
| [55] |
Bianchi S, Saglimbeni F, Frangipane G, et al. (2019) 3D dynamics of bacteria wall entrapment at a water–air interface. Soft Matter 15: 3397-3406. doi: 10.1039/C9SM00077A
|
| [56] |
Berg HC (1971) How to track bacteria. Rev Sci Instrum 42: 868-871. doi: 10.1063/1.1685246
|
| [57] |
Frymier PD, Ford RM, Berg HC, et al. (1995) Three-dimensional tracking of motile bacteria near a solid planar surface. P Natl Acad Sci USA 92: 6195-6199. doi: 10.1073/pnas.92.13.6195
|
| [58] | Bianchi S, Saglimbeni F, Di Leonardo R (2017) Holographic imaging reveals the mechanism of wall entrapment in swimming bacteria. Phys Rev X 7: 011010. |
| [59] |
Tokárová V, Perumal AS, Nayak M, et al. (2021) Patterns of bacterial motility in microfluidics-confining environments. P Natl Acad Sci USA 118: e2013925118. doi: 10.1073/pnas.2013925118
|
| [60] |
Conrad JC, Poling-Skutvik R (2018) Confined flow: Consequences and implications for bacteria and biofilms. Annu Rev Chem Biomol Eng 9: 175-200. doi: 10.1146/annurev-chembioeng-060817-084006
|
| [61] |
Drescher K, Dunkel J, Cisneros LH, et al. (2011) Fluid dynamics and noise in bacterial cell-cell and cell-surface scattering. P Natl Acad Sci USA 108: 10940-10945. doi: 10.1073/pnas.1019079108
|
| [62] |
Morse M, Huang A, Li G, et al. (2013) Molecular adsorption steers bacterial swimming at the air/water interface. Biophys J 105: 21-28. doi: 10.1016/j.bpj.2013.05.026
|
| [63] |
Mitchell JG, Kogure K (2006) Bacterial motility: links to the environment and a driving force for microbial physics. FEMS Microbiol Ecol 55: 3-16. doi: 10.1111/j.1574-6941.2005.00003.x
|
| [64] |
Donlan RM (2002) Biofilms: Microbial life on surfaces. Emerg Infect Dis J 8: 881. doi: 10.3201/eid0809.020063
|
| [65] |
Kearns DB (2010) A field guide to bacterial swarming motility. Nat Rev Microbiol 8: 634-644. doi: 10.1038/nrmicro2405
|
| [66] |
Guttenplan SB, Kearns DB (2013) Regulation of flagellar motility during biofilm formation. FEMS Microbiol Rev 37: 849-871. doi: 10.1111/1574-6976.12018
|

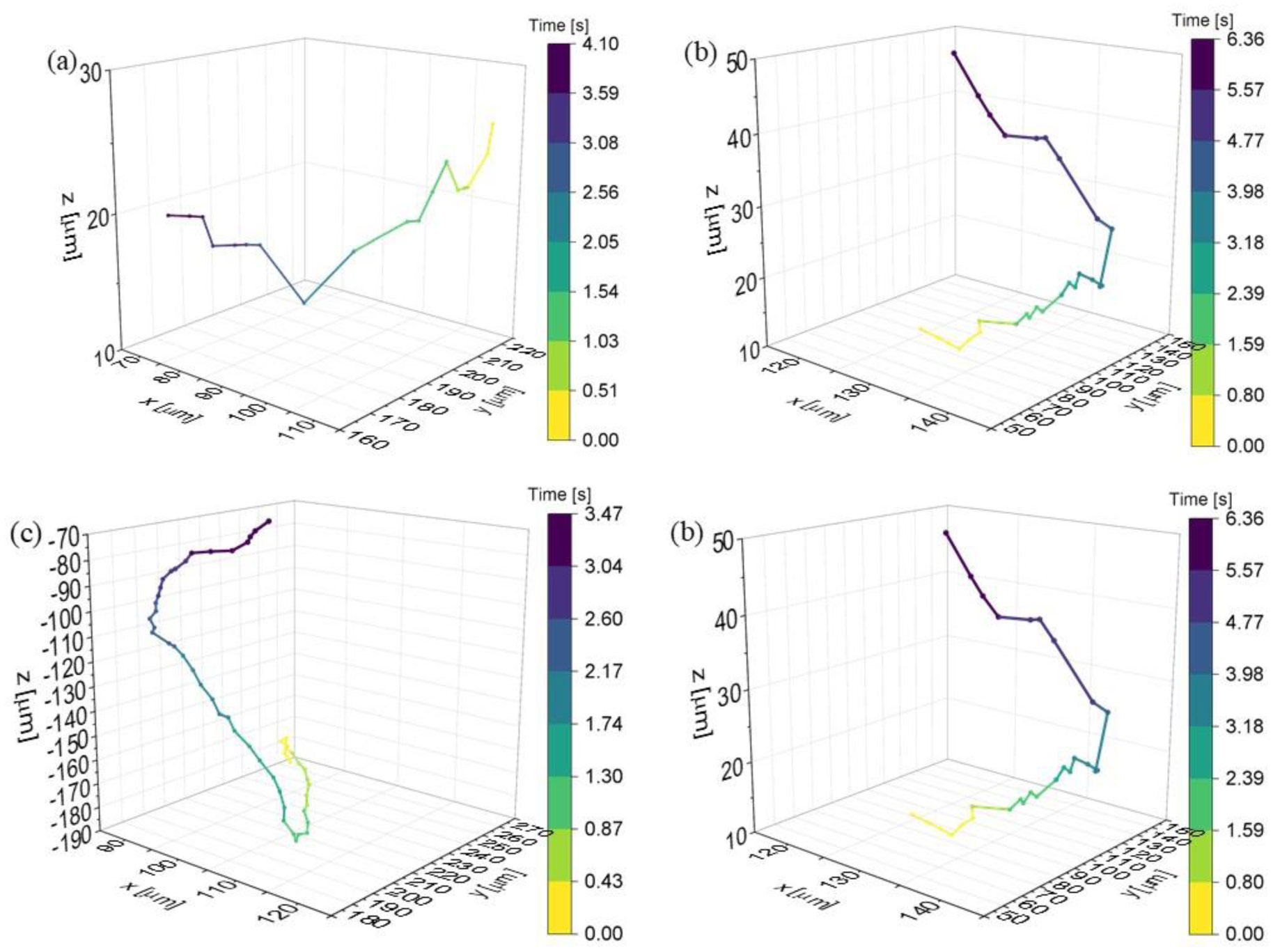
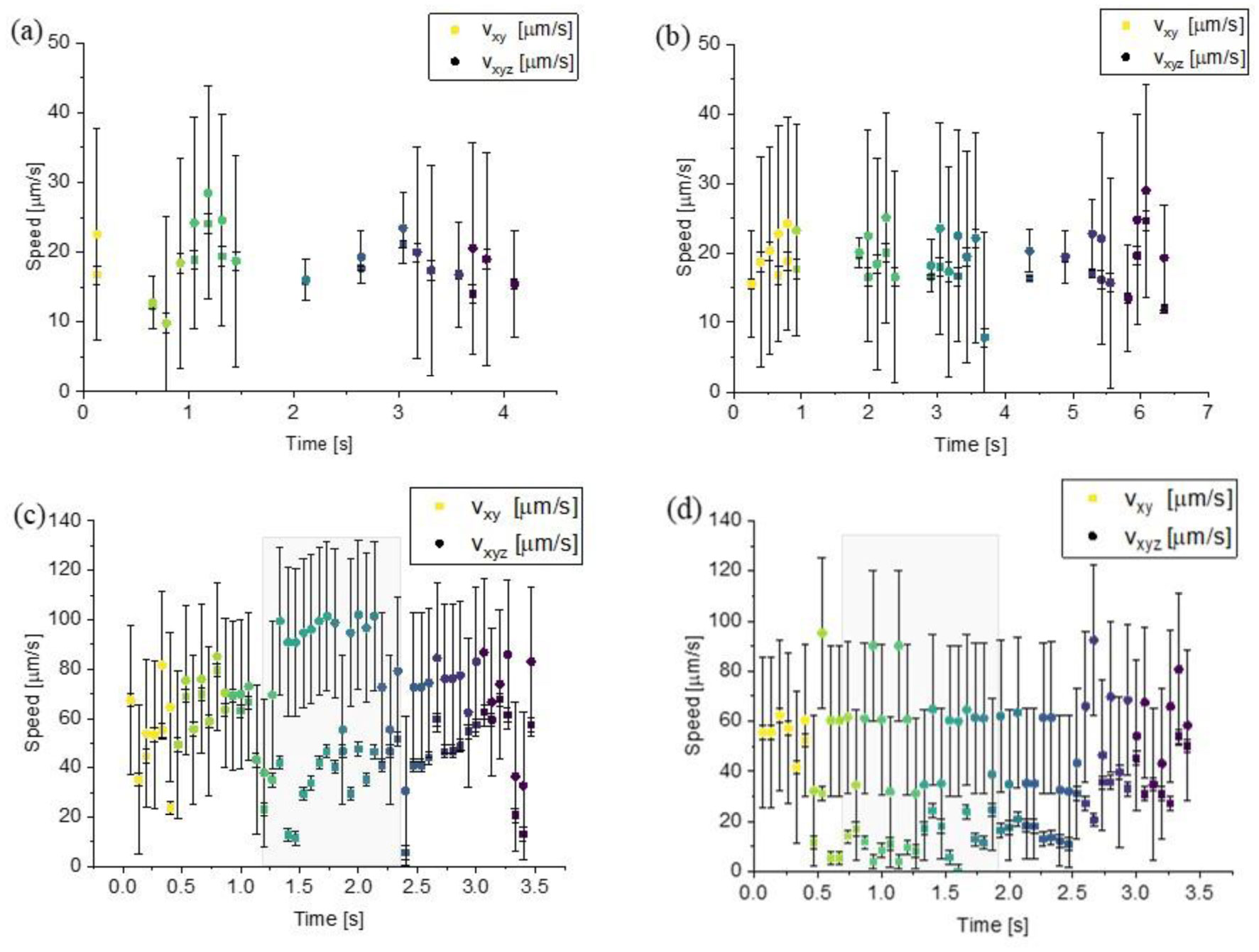

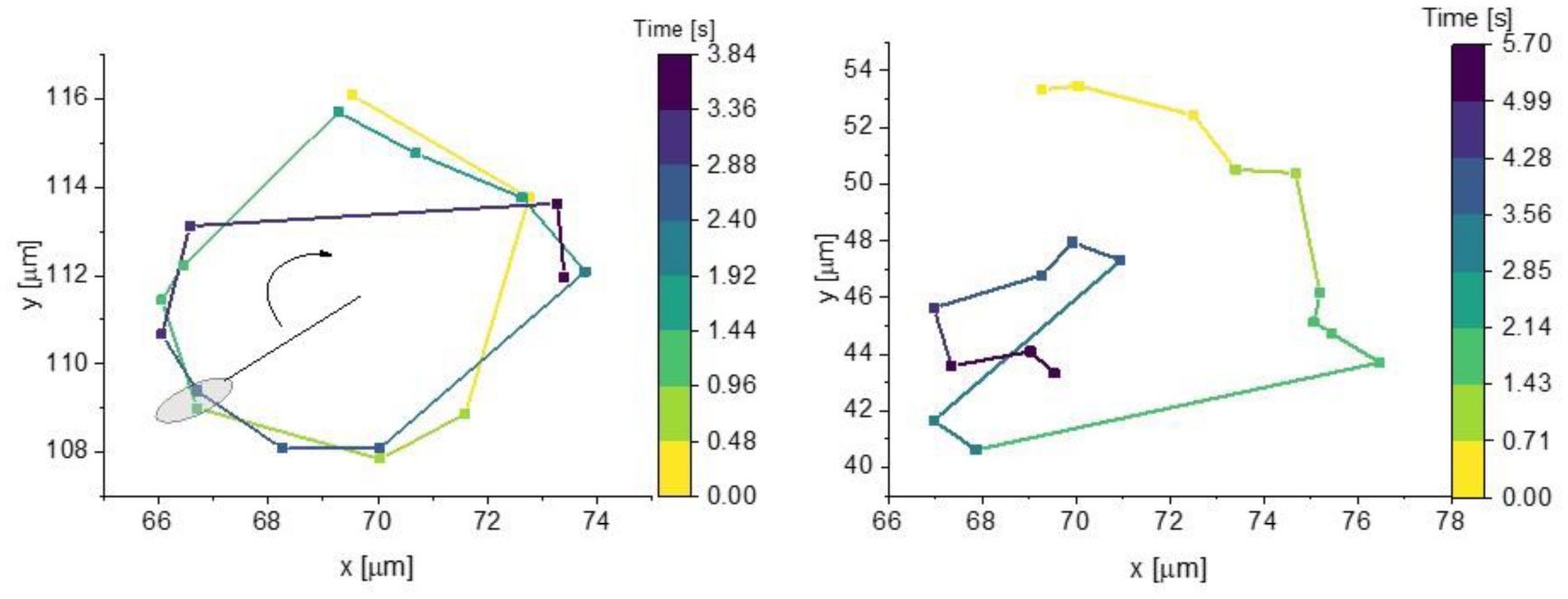
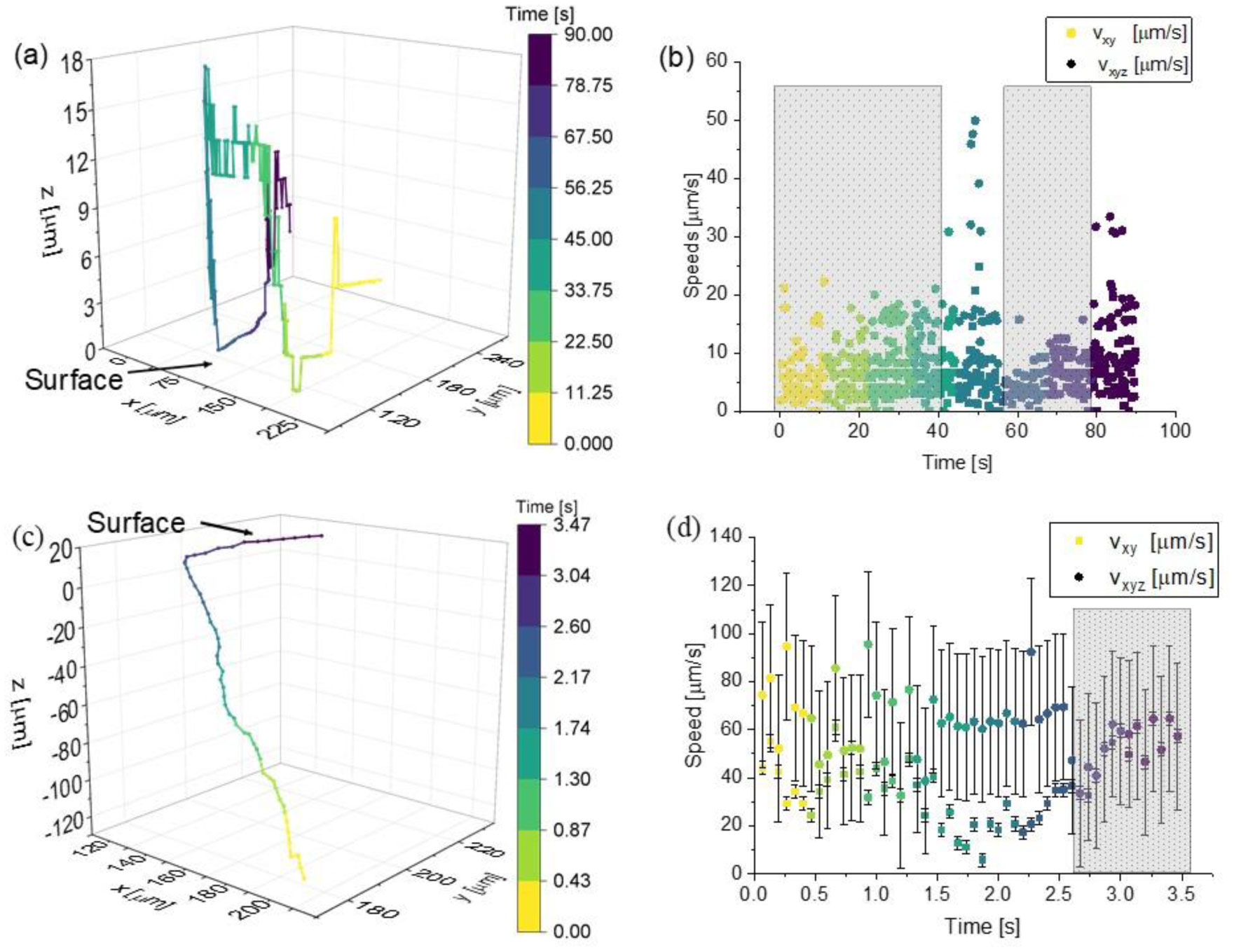
Figures(6) / Tables(1)

Jacqueline Acres, Jay Nadeau. 2D vs 3D tracking in bacterial motility analysis[J]. AIMS Biophysics, 2021, 8(4): 385-399. doi: 10.3934/biophy.2021030








 DownLoad:
DownLoad: