Citation: Yohei Matsunaga, Tsuyoshi Kawano. The C. elegans insulin-like peptides (ILPs)[J]. AIMS Biophysics, 2018, 5(4): 217-230. doi: 10.3934/biophy.2018.4.217

| [1] | Banting FG, Best CH, Collip JB, et al. (1922) Pancreatic extracts in the treatment of diabetes mellitus. Can Med Assoc J 12: 141–146. |
| [2] |
Ryle AP, Sanger F, Smith LF, et al. (1955) The disulphide bonds of insulin. Biochem J 60: 541–556. doi: 10.1042/bj0600541

|
| [3] | Dodson E, Harding MM, Hodgkin DC, et al. (1966) The crystal structure of insulin. 3. Evidence for a 2-fold axis in rhombohedral zinc insulin. J Mol Biol 16: 227–241. |
| [4] |
Mckern NM, Lawrence MC, Streltsov VA, et al. (2006) Structure of the insulin receptor ectodomain reveals a folded-over conformation. Nature 443: 218–221. doi: 10.1038/nature05106
|
| [5] | Haeusler RA, Mcgraw TE, Accili D (2018) Biochemical and cellular properties of insulin receptor signalling. Nat Rev Mol Cell Biol 19: 31–44. |
| [6] |
Vajdos FF, Ultsch M, Schaffer ML, et al. (2001) Crystal structure of humun insulin-like growth factor-1: Detergent binding inhibits binding protein interactions. Biochemistry 40: 11022–11029. doi: 10.1021/bi0109111
|
| [7] |
Hakuno F, Takahashi SI (2018) IGF1 receptor signaling pathways. J Mol Endocrinol 61: T69–T86. doi: 10.1530/JME-17-0311
|
| [8] | Fernandez AM, Torres-Aleman I (2012) The many faces of insulin-like peptide signaling in the brain. Nat Rev Neurosci 13: 225–239. |
| [9] |
Christie AE, Roncalli V, Lenz PH (2016) Diversity of insulin-like peptide signaling system proteins in Calanus finmarchicus (Crustacea; Copepoda)-possible contributors to seasonal pre-adult diapause. Gen Comp Endocrinol 236: 157–173. doi: 10.1016/j.ygcen.2016.07.016
|
| [10] | Lee Y, An SWA, Artan M, et al. (2015) Genes and pathways that influence longevity in Caenorhabditis elegans, In: Aging Mechanisms Mori N., Mook-Jung I, eds, 123–169. |
| [11] |
Giannakou ME, Partridge L (2007) Role of insulin-like signalling in Drosophila lifespan. Trends Biochem Sci 32: 180–188. doi: 10.1016/j.tibs.2007.02.007
|
| [12] |
Fontana L, Partridge L, Longo VD (2010) Extending healthy life span-from yeast to humans. Science 328: 321–326. doi: 10.1126/science.1172539
|
| [13] |
Kenyon CJ (2010) The genetics of ageing. Nature 464: 504–512. doi: 10.1038/nature08980
|
| [14] | Nassel DR, Kubrak OI, Liu Y, et al. (2013) Factors that regulate insulin producing cells and their output in Drosophila. Front Physiol 4: 252. |
| [15] | Grönke S, Partridge L, (2010) The functions of insulin-like peptides in insects, In: Clemmons D, Robinson I, Christen Y, eds., IGFs: Local repair and survival factors throughout life span, Research and Perspectives in Endocrine Interactions, Springer, Berlin, Heidelberg, 105–124. |
| [16] |
Zhang H, Liu J, Li CR, et al. (2009) Deletion of Drosophila insulin-like peptides causes growth defects and metabolic abnormalities. Proc Natl Acad Sci U S A 106: 19617–19622. doi: 10.1073/pnas.0905083106
|
| [17] |
Colombani J, Andersen DS, Leopold P (2012) Secreted peptide Dilp8 coordinates Drosophila tissue growth with developmental timing. Science 336: 582–585. doi: 10.1126/science.1216689
|
| [18] |
Abreu DAFD, Caballero A, Fardel P, et al. (2014) An insulin-to-insulin regulatory network orchestrates phenotypic specificity in development and physiology. PLoS Genet 10: e1004225. doi: 10.1371/journal.pgen.1004225
|
| [19] | Brenner S (1974) The genetics of Caenorhabditis elegans. Genetics 77: 91–94. |
| [20] | Consortium CES (1998) Genome sequence of the nematode C. elegans: A platform for investigating biology. Science 282: 2012–2018. |
| [21] |
Fire A, Xu S, Montgomery MK, et al. (1998) Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 391: 806–811. doi: 10.1038/35888
|
| [22] |
Zwaal RR, Broeks A, van Meurs J, et al. (1993) Target-selected gene inactivation in Caenorhabditis elegans by using a frozen transposon insertion mutant bank. Proc Natl Acad Sci U S A 90: 7431–7435. doi: 10.1073/pnas.90.16.7431
|
| [23] |
Klass MR (1983) A method for the isolation of longevity mutants in the nematode Caenorhabditis elegans and initial results. Mech Ageing Dev 22: 279–286. doi: 10.1016/0047-6374(83)90082-9
|
| [24] | Friedman DB, Johnson TE (1988) A mutation in the age-1 gene in Caenorhabditis elegans lengthens life and reduces hermaphrodite fertility. Genetics 118: 75–86. |
| [25] |
Friedman DB, Johnson TE (1988) Three mutants that extend both mean and maximum life span of the nematode, Caenorhabditis elegans, define the age-1 gene. J Gerontol Biol Sci 43: 102–109. doi: 10.1093/geronj/43.4.B102
|
| [26] |
Johnson TE (1990) Increased life-span of age-1 mutants in Caenorhabditis elegans and lower Gompertz rate of aging. Science 249: 908–912. doi: 10.1126/science.2392681
|
| [27] | Kenyon C, Chang J, Gensch E, et al. (1993) A C. elegans mutant that lives twice as long as wild type. Nature 366: 461–464. |
| [28] |
Kimura KD, Tissenbaum HA, Liu Y, et al. (1997) daf-2, an insulin receptor-like gene that regulates longevity and diapause in Caenorhabditis elegans. Science 277: 942–946. doi: 10.1126/science.277.5328.942
|
| [29] | Murakami S, Johnson TE (1996) A genetic pathway conferring life extension and resistance to UV stress in Caenorhabditis elegans. Genetics 143: 1207–1218. |
| [30] |
Cassada RC, Russell RL (1975) The dauerlarva, a post-embryonic developmental variant of the nematode Caenorhabditis elegans. Dev Biol 46: 326–342. doi: 10.1016/0012-1606(75)90109-8
|
| [31] |
Riddle DL, Swanson MM, Albert PS (1981) Interacting genes in nematode dauer larva formation. Nature 290: 668–671. doi: 10.1038/290668a0
|
| [32] | Vowels JJ, Thomas JH (1992) Genetic analysis of chemosensory control of dauer formation in Caenorhabditis elegans. Genetics 130: 105–123. |
| [33] | Gottlieb S, Ruvkun G (1994) daf-2, daf-16 and daf-23: Genetically interacting genes controlling Dauer formation in Caenorhabditis elegans. Genetics 137: 107–120. |
| [34] | Ogg S, Ruvkun G (1998) The C. elegans PTEN homolog, DAF-18, acts in the insulin receptor-like metabolic signaling pathway. Mol Cell 2: 887–893. |
| [35] |
Gil EB, Malone LE, Liu LX, et al. (1999) Regulation of the insulin-like developmental pathway of Caenorhabditis elegans by a homolog of the PTEN tumor suppressor gene. Proc Natl Acad Sci U S A 96: 2925–2930. doi: 10.1073/pnas.96.6.2925
|
| [36] |
Mihaylova VT, Borland CZ, Manjarrez L, et al. (1999) The PTEN tumor suppressor homolog in Caenorhabditis elegans regulates longevity and dauer formation in an insulin receptor-like signaling pathway. Proc Natl Acad Sci U S A 96: 7427–7432. doi: 10.1073/pnas.96.13.7427
|
| [37] |
Rouault JP, Kuwabara PE, Sinilnikova OM, et al. (1999) Regulation of dauer larva development in Caenorhabditis elegans by daf-18, a homologue of the tumour suppressor PTEN. Curr Biol 9: 329–332. doi: 10.1016/S0960-9822(99)80143-2
|
| [38] | Baugh LR, Sternberg PW (2006) DAF-16/FOXO regulates transcription of cki-1/Cip/Kip and repression of lin-4 during C. elegans L1 arrest. Curr Biol 16: 780–785. |
| [39] | Lithgow GJ, White TM, Hinerfeld DA, et al. (1994) Thermotolerance of a long-lived mutant of Caenorhabditis elegans. J Gerontol 49: 270–276. |
| [40] |
Lithgow GJ, White TM, Melov S, et al. (1995) Thermotolerance and extended life-span conferred by single-gene mutations and induced by thermal stress. Proc Natl Acad Sci U S A 92: 7540–7544. doi: 10.1073/pnas.92.16.7540
|
| [41] |
Babar P, Adamson C, Walker GA, et al. (1999) PI3-kinase inhibition induces dauer formation, thermotolerance and longevity in C. elegans. Neurobiol Aging 20: 513–519. doi: 10.1016/S0197-4580(99)00094-9
|
| [42] |
Walker GA, Walker DW, Lithgow GJ (1998) Genes that determine both thermotolerance and rate of aging in Caenorhabditis elegans. Ann N Y Acad Sci 851: 444–449. doi: 10.1111/j.1749-6632.1998.tb09022.x
|
| [43] |
Walker GA, White TM, McColl G, et al. (2001) Heat shock protein accumulation is upregulated in a long-lived mutant of Caenorhabditis elegans. J Gerontol A Biol Sci Med Sci 56: B281–B287. doi: 10.1093/gerona/56.7.B281
|
| [44] |
Hsu AL, Murphy CT, Kenyon C (2003) Regulation of aging and age-related disease by DAF-16 and heat-shock factor. Science 300: 1142–1145. doi: 10.1126/science.1083701
|
| [45] |
Honda Y, Honda S (1999) The daf-2 gene network for longevity regulates oxidative stress resistance and Mn-superoxide dismutase gene expression in Caenorhabditis elegans. FASEB J 13: 1385–1393. doi: 10.1096/fasebj.13.11.1385
|
| [46] |
Honda Y, Honda S (2002) Oxidative stress and life span determination in the nematode Caenorhabditis elegans. Ann N Y Acad Sci 959: 466–474. doi: 10.1111/j.1749-6632.2002.tb02117.x
|
| [47] | Scott BA, Avidan MS, Crowder CM (2002) Regulation of hypoxic death in C. elegans by the insulin/IGF receptor homolog DAF-2. Science 296: 2388–2391. |
| [48] | Lamitina ST, Strange K (2005) Transcriptional targets of DAF-16 insulin signaling pathway protect C. elegans from extreme hypertonic stress. Am J Physiol -Cell Ph 288: C467–C474. |
| [49] |
Barsyte D, Lovejoy DA, Lithgow GJ (2001) Longevity and heavy metal resistance in daf-2 and age-1 long-lived mutants of Caenorhabditis elegans. FASEB J 15: 627–634. doi: 10.1096/fj.99-0966com
|
| [50] |
Morley JF, Morimoto RI (2004) Regulation of longevity in Caenorhabditis elegans by heat shock factor and molecular chaperones. Mol Biol Cell 15: 657–664. doi: 10.1091/mbc.e03-07-0532
|
| [51] |
Cohen E, Bieschke J, Perciavalle RM, et al. (2006) Opposing activities protect against age-onset proteotoxicity. Science 313: 1604–1610. doi: 10.1126/science.1124646
|
| [52] |
Keowkase R, Aboukhatwa M, Luo Y (2010) Fluoxetine protects against amyloid-beta toxicity, in part via daf-16 mediated cell signaling pathway, in Caenorhabditis elegans. Neuropharmacology 59: 358–365. doi: 10.1016/j.neuropharm.2010.04.008
|
| [53] | Ching TT, Chiang WC, Chen CS, et al. (2011) Celecoxib extends C. elegans lifespan via inhibition of insulin-like signaling but not cyclooxygenase-2 activity. Aging Cell 10: 506–519. |
| [54] | Teixeira-Castro A, Ailion M, Jalles A, et al. (2011) Neuron-specific proteotoxicity of mutant ataxin-3 in C. elegans: Rescue by the DAF-16 and HSF-1 pathways. Hum Mol Genet 20: 2996–3009. |
| [55] |
Zhang T, Hwang HY, Hao H, et al. (2012) Caenorhabditis elegans RNA-processing protein TDP-1 regulates protein homeostasis and life span. J Biol Chem 287: 8371–8382. doi: 10.1074/jbc.M111.311977
|
| [56] |
Nagasawa H, Kataoka H, Isogai A, et al. (1984) Amino-terminal amino acid sequence of the silkworm prothoracicotropic hormone: Homology with insulin. Science 226: 1344–1345. doi: 10.1126/science.226.4680.1344
|
| [57] |
Smit AB, Vreugdenhil E, Ebberink RH, et al. (1988) Growth-controlling molluscan neurons produce the precursor of an insulin-related peptide. Nature 331: 535–538. doi: 10.1038/331535a0
|
| [58] |
Lagueux M, Lwoff L, Meister M, et al. (1990) cDNAs from neurosecretory cells of brains of Locusta migratoria (Insecta, Orthoptera) encoding a novel member of the superfamily of insulins. Eur J Biochem 187: 249–254. doi: 10.1111/j.1432-1033.1990.tb15302.x
|
| [59] |
Chandler JC, Aizen J, Elizur A, et al. (2015) Discovery of a novel insulin-like peptide and insulin binding proteins in the Eastern rock lobster Sagmariasus verreauxi. Gen Comp Endocr 215: 76–87. doi: 10.1016/j.ygcen.2014.08.018
|
| [60] |
Duret L, Guex N, Peitsch MC, et al. (1998) New insulin-like proteins with atypical disulfide bond pattern characterized in Caenorhabditis elegans by comparative sequence analysis and homology modeling. Genome Res 8: 348–353. doi: 10.1101/gr.8.4.348
|
| [61] | Pierce SB, Costa M, Wisotzkey R, et al. (2001) Regulation of DAF-2 receptor signaling by human insulin and ins-1, a member of the unusually large and diverse C. elegans insulin gene family. Genes Dev 15: 672–686. |
| [62] | Li W, Kennedy SG, Ruvkun G (2003) daf-28 encodes a C. elegans insulin superfamily member that is regulated by environmental cues and acts in the DAF-2 signaling pathway. Genes Dev 17: 844–858. |
| [63] |
Husson SJ, Mertens I, Janssen T, et al. (2007) Neuropeptidergic signaling in the nematode Caenorhabditis elegans. Prog Neurobiol 82: 33–55. doi: 10.1016/j.pneurobio.2007.01.006
|
| [64] |
Kawano T, Ito Y, Ishiguro M, et al. (2000) Molecular cloning and characterization of a new insulin/IGF-like peptide of the nematode Caenorhabditis elegans. Biochem Biophys Res Commun 273: 431–436. doi: 10.1006/bbrc.2000.2971
|
| [65] |
Matsunaga Y, Gengyo-Ando K, Mitani S, et al. (2012) Physiological function, expression pattern, and transcriptional regulation of a Caenorhabditis elegans insulin-like peptide, INS-18. Biochem Biophys Res Commun 423: 478–483. doi: 10.1016/j.bbrc.2012.05.145
|
| [66] | Wang X, Wang X, Wang D, et al. (2010) [Expression changes of age-related genes in different aging stages of Caenorhabiditis elegans and the regulating effects of Chuanxiong extract]. China J Chin Mater Med 35: 1599–1602. |
| [67] |
Kawano T, Nagatomo R, Kimura Y, et al. (2006) Disruption of ins-11, a Caenorhabditis elegans insulin-like gene, and phenotypic analyses of the gene-disrupted animal. Biosci Biotechnol Biochem 70: 3084–3087. doi: 10.1271/bbb.60472
|
| [68] |
Morris JZ, Tissenbaum HA, Ruvkun G (1996) A phosphatidylinositol-3-OH kinase family member regulating longevity and diapause in Caenorhabditis elegans. Nature 382: 536–539. doi: 10.1038/382536a0
|
| [69] |
Paradis S, Ailion M, Toker A, et al. (1999) A PDK1 homolog is necessary and sufficient to transduce AGE-1 PI3 kinase signals that regulate diapause in Caenorhabditis elegans. Genes Dev 13: 1438–1452. doi: 10.1101/gad.13.11.1438
|
| [70] |
Paradis S, Ruvkun G (1998) Caenorhabditis elegans Akt/PKB transduces insulin receptor-like signals from AGE-1 PI3 kinase to the DAF-16 transcription factor. Genes Dev 12: 2488–2498. doi: 10.1101/gad.12.16.2488
|
| [71] |
Ogg S, Paradis S, Gottlieb S, et al. (1997) The Fork head transcription factor DAF-16 transduces insulin-like metabolic and longevity signals in C. elegans. Nature 389: 994–999. doi: 10.1038/40194
|
| [72] |
Henderson ST, Johnson TE (2001) daf-16 integrates developmental and environmental inputs to mediate aging in the nematode Caenorhabditis elegans. Curr Biol 11: 1975–1980. doi: 10.1016/S0960-9822(01)00594-2
|
| [73] | Lee RY, Hench J, Ruvkun G (2001) Regulation of C. elegans DAF-16 and its human ortholog FKHRL1 by the daf-2 insulin-like signaling pathway. Curr Biol 11: 1950–1957. |
| [74] | Malone EA, Inoue T, Thomas JH (1996) Genetic analysis of the roles of daf-28 and age-1 in regulating Caenorhabditis elegans dauer formation. Genetics 143: 1193–1205. |
| [75] |
Murphy CT, Mccarroll SA, Bargmann CI, et al. (2003) Genes that act downstream of DAF-16 to influence the lifespan of Caenorhabditis elegans. Nature 424: 277–283. doi: 10.1038/nature01789
|
| [76] |
Matsunaga Y, Nakajima K, Gengyo-Ando K, et al. (2012) A Caenorhabditis elegans insulin-like peptide, INS-17: Its physiological function and expression pattern. Biosci Biotechnol Biochem 76: 2168–2172. doi: 10.1271/bbb.120540
|
| [77] |
Matsunaga Y, Matsukawa T, Iwasaki T, et al. (2018) Comparison of physiological functions of antagonistic insulin-like peptides, INS-23 and INS-18, in Caenorhabditis elegans. Biosci Biotechnol Biochem 82: 90–96. doi: 10.1080/09168451.2017.1415749
|
| [78] |
Cornils A, Gloeck M, Chen Z, et al. (2011) Specific insulin-like peptides encode sensory information to regulate distinct developmental processes. Development 138: 1183–1193. doi: 10.1242/dev.060905
|
| [79] |
Murphy CT, Lee SJ, Kenyon C (2007) Tissue entrainment by feedback regulation of insulin gene expression in the endoderm of Caenorhabditis elegans. Proc Natl Acad Sci U S A 104: 19046–19050. doi: 10.1073/pnas.0709613104
|
| [80] |
Tomioka M, Adachi T, Suzuki H, et al. (2006) The insulin/PI 3-kinase pathway regulates salt chemotaxis learning in Caenorhabditis elegans. Neuron 51: 613–625. doi: 10.1016/j.neuron.2006.07.024
|
| [81] |
Kodama E, Kuhara A, Mohri-Shiomi A, et al. (2006) Insulin-like signaling and the neural circuit for integrative behavior in C. elegans. Genes Dev 20: 2955–2960. doi: 10.1101/gad.1479906
|
| [82] |
Lin CH, Tomioka M, Pereira S, et al. (2010) Insulin signaling plays a dual role in Caenorhabditis elegans memory acquisition and memory retrieval. J Neurosci 30: 8001–8011. doi: 10.1523/JNEUROSCI.4636-09.2010
|
| [83] |
Chen Z, Hendricks M, Cornils A, et al. (2013) Two insulin-like peptides antagonistically regulate aversive olfactory learning in C. elegans. Neuron 77: 572–585. doi: 10.1016/j.neuron.2012.11.025
|
| [84] | Michaelson D, Korta DZ, Capua Y, et al. (2010) Insulin signaling promotes germline proliferation in C. elegans. Development 137: 671–680. |
| [85] | Ritter AD, Shen Y, Fuxman BJ, et al. (2013) Complex expression dynamics and robustness in C. elegans insulin networks. Genome Res 23: 954–965. |
| [86] | Matsunaga Y, Iwasaki T, Kawano T (2017) Diverse insulin-like peptides in Caenorhabditis elegans. Int Biol Rev , 1. |
| [87] |
Hua QX, Nakagawa SH, Wilken J, et al. (2003) A divergent INS protein in Caenorhabditis elegans structurally resembles human insulin and activates the human insulin receptor. Genes Dev 17: 826–831. doi: 10.1101/gad.1058003
|
| [88] |
Kao G, Nordenson C, Still M, et al. (2007) ASNA-1 positively regulates insulin secretion in C. elegans and mammalian cells. Cell 128: 577–587. doi: 10.1016/j.cell.2006.12.031
|
| [89] |
Matsunaga Y, Honda Y, Honda S, et al. (2016) Diapause is associated with a change in the polarity of secretion of insulin-like peptides. Nat Commun 7: 10573. doi: 10.1038/ncomms10573
|
| [90] |
Clark ME, Kelner GS, Turbeville LA, et al. (2000) ADAMTS9, a novel member of the ADAM-TS/ metallospondin gene family. Genomics 67: 343–350. doi: 10.1006/geno.2000.6246
|
| [91] |
Somerville RP, Longpre JM, Jungers KA, et al. (2003) Characterization of ADAMTS-9 and ADAMTS-20 as a distinct ADAMTS subfamily related to Caenorhabditis elegans GON-1. J Biol Chem 278: 9503–9513. doi: 10.1074/jbc.M211009200
|
| [92] | Yoshina S, Mitani S (2015) Loss of C. elegans GON-1, an ADAMTS9 Homolog, Decreases Secretion Resulting in Altered Lifespan and Dauer Formation. PLoS One 10: e0133966. |
| [93] |
Suckale J, Solimena M (2010) The insulin secretory granule as a signaling hub. Trends Endocrin Met 21: 599–609. doi: 10.1016/j.tem.2010.06.003
|
| [94] |
Drucker DJ, Philippe J, Mojsov S, et al. (1987) Glucagon-like peptide I stimulates insulin gene expression and increases cyclic AMP levels in a rat islet cell line. Proc Natl Acad Sci U S A 84: 3434–3438. doi: 10.1073/pnas.84.10.3434
|
| [95] |
Fehmann HC, Habener JF (1992) Galanin inhibits proinsulin gene expression stimulated by the insulinotropic hormone glucagon-like peptide-I (7-37) in mouse insulinoma beta TC-1 cells. Endocrinology 130: 2890–2896. doi: 10.1210/endo.130.5.1374016
|
| [96] |
Koranyi L, James DE, Kraegen EW, et al. (1992) Feedback inhibition of insulin gene expression by insulin. J Clin Invest 89: 432–436. doi: 10.1172/JCI115602
|
| [97] | Shaw WM, Luo S, Landis J, et al. (2007) The C. elegans TGF-beta Dauer pathway regulates longevity via insulin signaling. Curr Biol 17: 1635–1645. |
| [98] |
Hung WL, Wang Y, Chitturi J, et al. (2014) A Caenorhabditis elegans developmental decision requires insulin signaling-mediated neuron-intestine communication. Development 141: 1767–1779. doi: 10.1242/dev.103846
|
| [99] |
Liu T, Zimmerman KK, Patterson GI (2004) Regulation of signaling genes by TGFbeta during entry into dauer diapause in C. elegans. BMC Dev Biol 4: 11. doi: 10.1186/1471-213X-4-11
|
| [100] |
Narasimhan SD, Yen K, Bansal A, et al. (2011) PDP-1 links the TGF-beta and IIS pathways to regulate longevity, development, and metabolism. PLoS Genet 7: e1001377. doi: 10.1371/journal.pgen.1001377
|
| [101] |
Dlakic M (2002) A new family of putative insulin receptor-like proteins in C. elegans. Curr Biol 12: R155–R157. doi: 10.1016/S0960-9822(02)00729-7
|
| [102] | Tomioka M, Naito Y, Kuroyanagi H, et al. (2016) Splicing factors control C. elegans behavioural learning in a single neuron by producing DAF-2c receptor. Nat Commun 7: 11645. |
| [103] |
Bulger DA, Fukushige T, Yun S, et al. (2017) Caenorhabditis elegans DAF-2 as a model for human insulin receptoropathies. G3 Genesgenetics 7: 257–268. doi: 10.1534/g3.116.037184
|
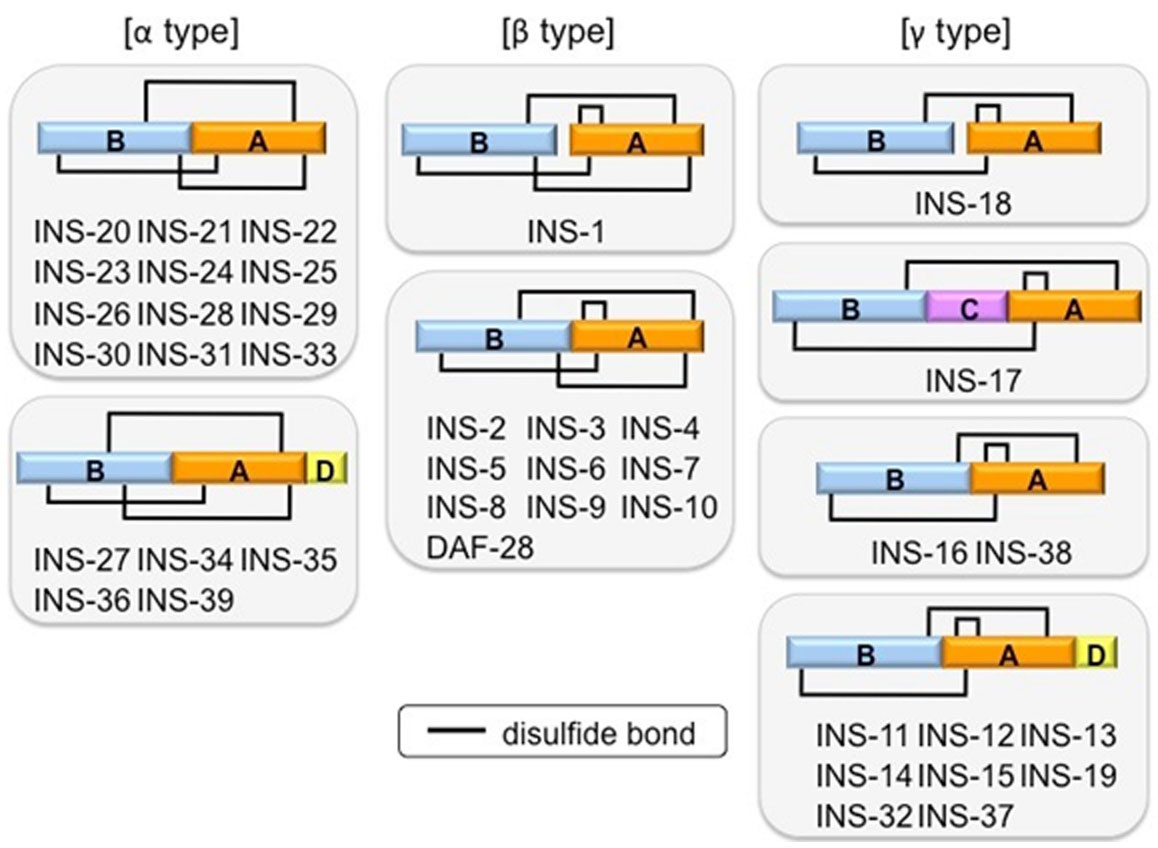
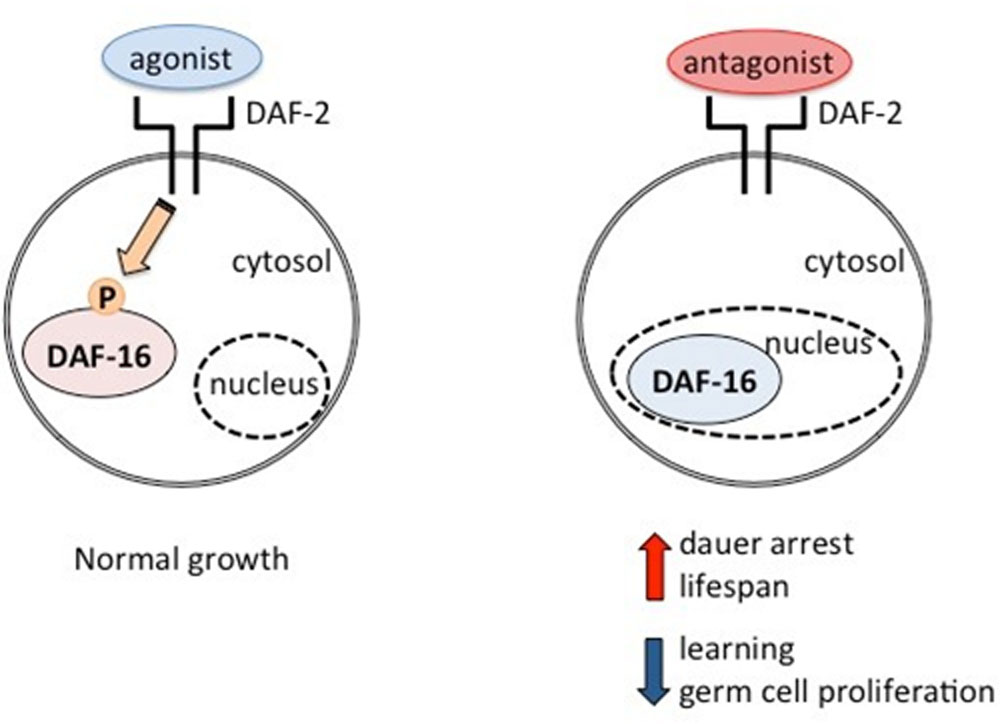

Figures(3)

Yohei Matsunaga, Tsuyoshi Kawano. The C. elegans insulin-like peptides (ILPs)[J]. AIMS Biophysics, 2018, 5(4): 217-230. doi: 10.3934/biophy.2018.4.217







 DownLoad:
DownLoad: