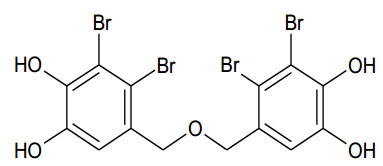
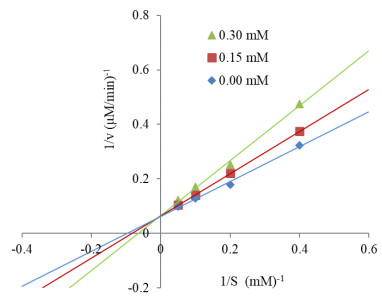
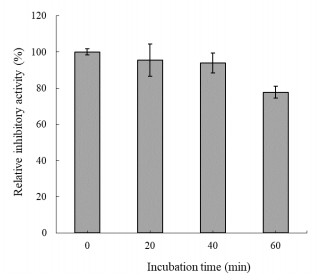
The human gastrointestinal (GI) tract plays an important role in disease and health. The overexpression of β-glucuronidase, an enzyme produced in the gastrointestinal tract, has been found to cause the retoxification of potentially damaging substances already detoxified by liver glucuronidation. Thus, β-glucuronidase has been implicated in a number of diseases including colon cancer, liver disease and Crohn's disease. This study investigated β-glucuronidase inhibitory activity of a bromophenol purified from the red alga Grateloupia lancifolia. Using an assay-guided fractionation technique, a bromophenol, bis(2, 3-dibromo-4, 5-dihydroxybenzyl) ether, a bromophenol, was isolated from the red alga G. lancifolia. Its structure was elucidated on the basis of extensive 1D- and 2D-NMR studies, high resolution EI-MS analysis and comparisons with literature data. Bis(2, 3-dibromo-4, 5-dihydroxybenzyl) ether showed potent β-glucuronidase inhibitory activity. The IC50 and KI values of bromophenol against E. coli β-glucuronidase were 0.54 and 0.53 mM, respectively. This bromophenol inhibited β-glucuronidase competitively and was stable at pH 2 up to 40 min at 37 ℃.
Citation: The Han Nguyen, Patrick Achiever Blamo Jr, Xiaoyong Liu, Thi Van Anh Tran, Sang Moo Kim. β-Glucuronidase inhibitory activity of bromophenol isolated from red alga Grateloupia lancifolia[J]. AIMS Agriculture and Food, 2021, 6(2): 551-559. doi: 10.3934/agrfood.2021032
The human gastrointestinal (GI) tract plays an important role in disease and health. The overexpression of β-glucuronidase, an enzyme produced in the gastrointestinal tract, has been found to cause the retoxification of potentially damaging substances already detoxified by liver glucuronidation. Thus, β-glucuronidase has been implicated in a number of diseases including colon cancer, liver disease and Crohn's disease. This study investigated β-glucuronidase inhibitory activity of a bromophenol purified from the red alga Grateloupia lancifolia. Using an assay-guided fractionation technique, a bromophenol, bis(2, 3-dibromo-4, 5-dihydroxybenzyl) ether, a bromophenol, was isolated from the red alga G. lancifolia. Its structure was elucidated on the basis of extensive 1D- and 2D-NMR studies, high resolution EI-MS analysis and comparisons with literature data. Bis(2, 3-dibromo-4, 5-dihydroxybenzyl) ether showed potent β-glucuronidase inhibitory activity. The IC50 and KI values of bromophenol against E. coli β-glucuronidase were 0.54 and 0.53 mM, respectively. This bromophenol inhibited β-glucuronidase competitively and was stable at pH 2 up to 40 min at 37 ℃.

| [1] |
Thursby E, Juge N (2017) Introduction to the human gut microbiota. Biochemical J 474: 1823-1836. doi: 10.1042/BCJ20160510

|
| [2] |
Li CQ, Zhang JY, Xu H, et al. (2019) Effect of Dahuang Danpi Decoction on Lactobacillus bulgaricus growth and metabolism: In vitro study. Medicine 98: e13941. doi: 10.1097/MD.0000000000013941
|
| [3] |
Li JH, Jia HJ, Cai XH, et al. (2014) An integrated catalog of reference genes in the human gut microbiome. Nat Biotechnol 32: 834-841. doi: 10.1038/nbt.2942
|
| [4] |
Rowland I, Gibson G, Heinken A, et al. (2018) Gut microbiota functions: Metabolism of nutrients and other food components. Eur J Nutr 57: 1-24. doi: 10.1007/s00394-017-1445-8
|
| [5] | Kim KY, Choi KS, Kurihara H, et al. (2008) β-Glucuronidase inhibitory activity of bromophenols purified from Grateloupia elliptica. Food Sci Biotechnol 17: 1110-1114. |
| [6] |
Gloux K, Anba-Mondoloni J (2016) Unique β-glucuronidase locus in gut microbiomes of Crohn's disease patients and unaffected first-degree relatives. PLoS One 11: e0148291. doi: 10.1371/journal.pone.0148291
|
| [7] | Sekikawa C, Kurihara H, Goto K, et al. (2002) Inhibition of β-glucuronidase by extracts of Chondria crassicaulis. Bull Fish Sci Hokkaido Univ 53: 27-30. |
| [8] |
Kim DH, Jin YH (2001) Intestinal bacterial β-glucuronidase activity of patients with colon cancer. Arch Pharm Res 24: 564-567. doi: 10.1007/BF02975166
|
| [9] |
Kim DH, Shim SB, Kim NJ, et al. (1999) Beta-glucuronidase inhibitory activity and hepatoprotective effect of Ganoderma lucidum. Biol Pharm Bull 2: 162-164. doi: 10.1248/bpb.22.162
|
| [10] |
Karak S, Nag G, De B (2017) Metabolic profile and β-glucuronidase inhibitory property of three species of swertia. Rev Bras Farmacogn 27: 105-111. doi: 10.1016/j.bjp.2016.07.007
|
| [11] |
Chakraborty K, Maneesh A, Makkar F (2017) Antioxidant activity of brown seaweeds. J Aquat Food Prod T 26: 406-419. doi: 10.1080/10498850.2016.1201711
|
| [12] |
El Shafay SM, Ali SS, El-Sheekh MM (2016) Antimicrobial activity of some seaweeds species from red sea, against multidrug resistant bacteria. Egypt J Aquat Res 42: 65-74. doi: 10.1016/j.ejar.2015.11.006
|
| [13] |
Usoltseva RV, Anastyuk SD, Ishina IA, et al. (2018) Structural characteristics and anticancer activity in vitro of fucoidan from brown alga Padina boryana. Carbohydr Polym 184: 260-268. doi: 10.1016/j.carbpol.2017.12.071
|
| [14] |
Apostolidis E, Karayannakidis PD, Kwon YI, et al. (2011) Seasonal variation of phenolic antioxidant-mediated α-glucosidase inhibition of Ascophyllum nodosum. Plant Foods Hum Nutr 66: 313-319. doi: 10.1007/s11130-011-0250-4
|
| [15] |
Yermakova SP, Sova VV, Zvyagintseva TN (2002) Brown seaweed protein as an inhibitor of marine mollusk endo-(1→3)-β-D-glucanases. Carbohydr Res 337: 229-237. doi: 10.1016/S0008-6215(01)00304-4
|
| [16] |
Islam MR, Arslan I, Attia J, et al. (2013) Is serum zinc level associated with prediabetes and diabetes?: A cross-sectional study from Bangladesh. PLoS One 8: e61776. doi: 10.1371/journal.pone.0061776
|
| [17] |
Nguyen TH, Kim SM (2012) Antioxidative, anticholinesterase and antityrosinase activities of the red alga Polyopes lancifolia extracts. Afr J Biotechnol 11: 9457-9467. doi: 10.5897/AJB12.1166
|
| [18] | Kim KY, Nguyen TH, Kurihara H, et al. (2010) α-Glucosidase inhibitory activity of bromophenol purified from the red alga Polyopes lancifolia. J Food Sci 75: H145-H150. |
| [19] | De Clerck O, Gavio B, Fredericq S, et al. (2005) Systematic of Grateloupia filicina (Halymeniaceae, Rhodophyta), based on RBCL sequence analyses and morphological evidence, including the reinstatement of G. minima and the description of G. Capensis SP. NOV.1: Systematic of Grateloupia filicina. J Phycol 41: 391-410. |
| [20] |
Lineweaver H, Burk D (1934) The determination of enzyme dissociation constants. J Am Chem Soc 56: 658-666. doi: 10.1021/ja01318a036
|
| [21] | Suffness M, Newman DJ, Snader K (1989) Bioorganic marine chemistry. In: Scheuer PJ (Ed), Discovery and development of antineoplastic agents from natural sources. Berlin: Springer-Verlag, 131-168. |
| [22] |
Fan X, Xu NJ, Shi JG (2003) Bromophenols from red alga Rhodomela confervoides. J Nat Prod 66: 455-458. doi: 10.1021/np020528c
|
| [23] |
Imran S, Taha M, Ismail NH, et al. (2015) Synthesis, biological evaluation, and docking studies of novel thiourea derivatives of bisindolylmethane as carbonic anhydrase Ⅱ inhibitor. Bioorg Chem 62: 83-93. doi: 10.1016/j.bioorg.2015.08.001
|
| [24] |
Mekinić IG, Skroza D, Šimat V, et al. (2019) Phenolic content of brown algae (Pheophyceae) species: Extraction, identification, and quantification. Biomolecules 9: 244. doi: 10.3390/biom9060244
|
| [25] |
De Neve N, Vlaeminck B, Gadeyne F, et al. (2018) Protection of emulsified polyunsaturated fatty acids against in vitro ruminal biohydrogenation by polyphenol oxidase: Characterization of the cross-linked emulsion. Anim Feed Sci Tech 246: 91-103. doi: 10.1016/j.anifeedsci.2018.10.004
|
| [26] |
Le P, Zhao J, Franzen S (2014) Correlation of heme binding affinity and enzyme kinetics of dehaloperoxidase. Biochemistry 53: 6863-6877. doi: 10.1021/bi5005975
|
| [27] |
Hong NS, Petrović D, Lee R, et al. (2018) The evolution of multiple active site configurations in a designed enzyme. Nat Commun 9: 3900. doi: 10.1038/s41467-018-06305-y
|
| [28] |
Masola V, Bellin G, Gambaro G, et al. (2018) Heparanase: A multitasking protein involved in extracellular matrix (ECM) remodeling and intracellular events. Cells 7: 236. doi: 10.3390/cells7120236
|
| [29] |
Meirovitz A, Goldberg R, Binder A, et al. (2013) Heparanase in inflammation and inflammation-associated cancer. FEBS J 280: 2307-2319. doi: 10.1111/febs.12184
|
| [30] | Bae EA, Kim DH, Han MJ (2001) Effect of Lentinus edodes water extract on some enzymes of mouse intestinal bacteria. Korean J Food Sci Technol 33: 142-145. |
| [31] |
Gudiel-Urbano M, Goni I (2002) Effect of edible seaweeds (Undaria pinnatifida and Porphyra ternera) on the metabolic activities of intestinal microflora in rats. Nutr Res 22: 323-331. doi: 10.1016/S0271-5317(01)00383-9
|
| [32] |
Satoh T, Igarashi A, Tanno M, et al. (2018) Inhibitory effects of baicalein derived from Japanese traditional herbal medicine on SN-38 glucuronidation. J Pharm Pharm Sci 21: 195-206. doi: 10.18433/jpps29783
|
| [33] | Guo YW, Chen YH, Chiu WC, et al. (2013) Soy saponins meditate the progression of colon cancer in rats by inhibiting the activity of β-glucuronidase and the number of aberrant Crypt foci but not cyclooxygenase-2 activity. ISRN Oncol 645817. |
Figures(3) / Tables(2)

The Han Nguyen, Patrick Achiever Blamo Jr, Xiaoyong Liu, Thi Van Anh Tran, Sang Moo Kim. β-Glucuronidase inhibitory activity of bromophenol isolated from red alga Grateloupia lancifolia[J]. AIMS Agriculture and Food, 2021, 6(2): 551-559. doi: 10.3934/agrfood.2021032







 DownLoad:
DownLoad: