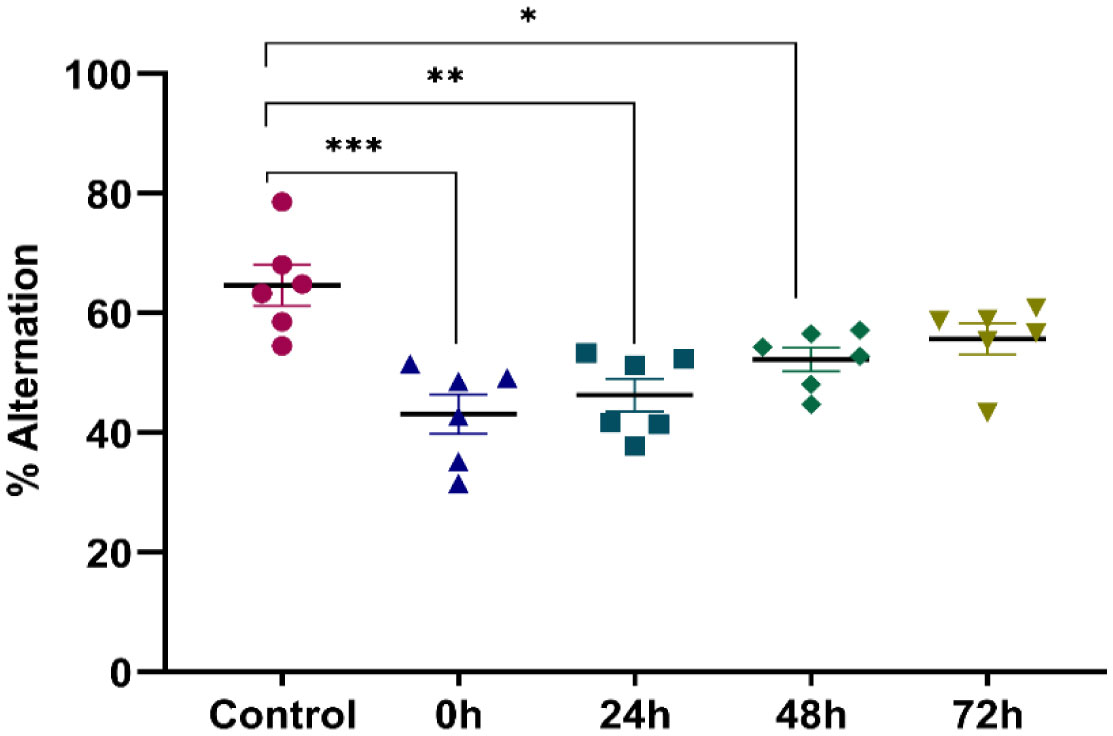
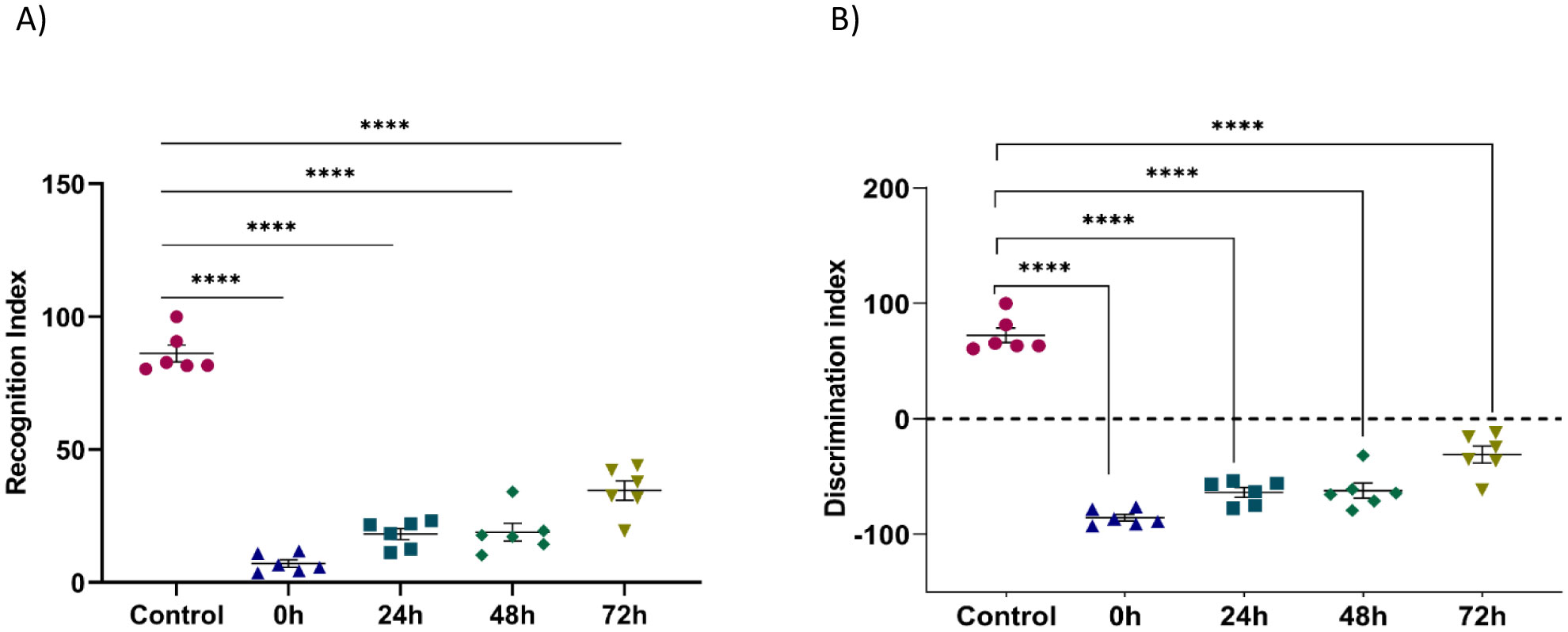
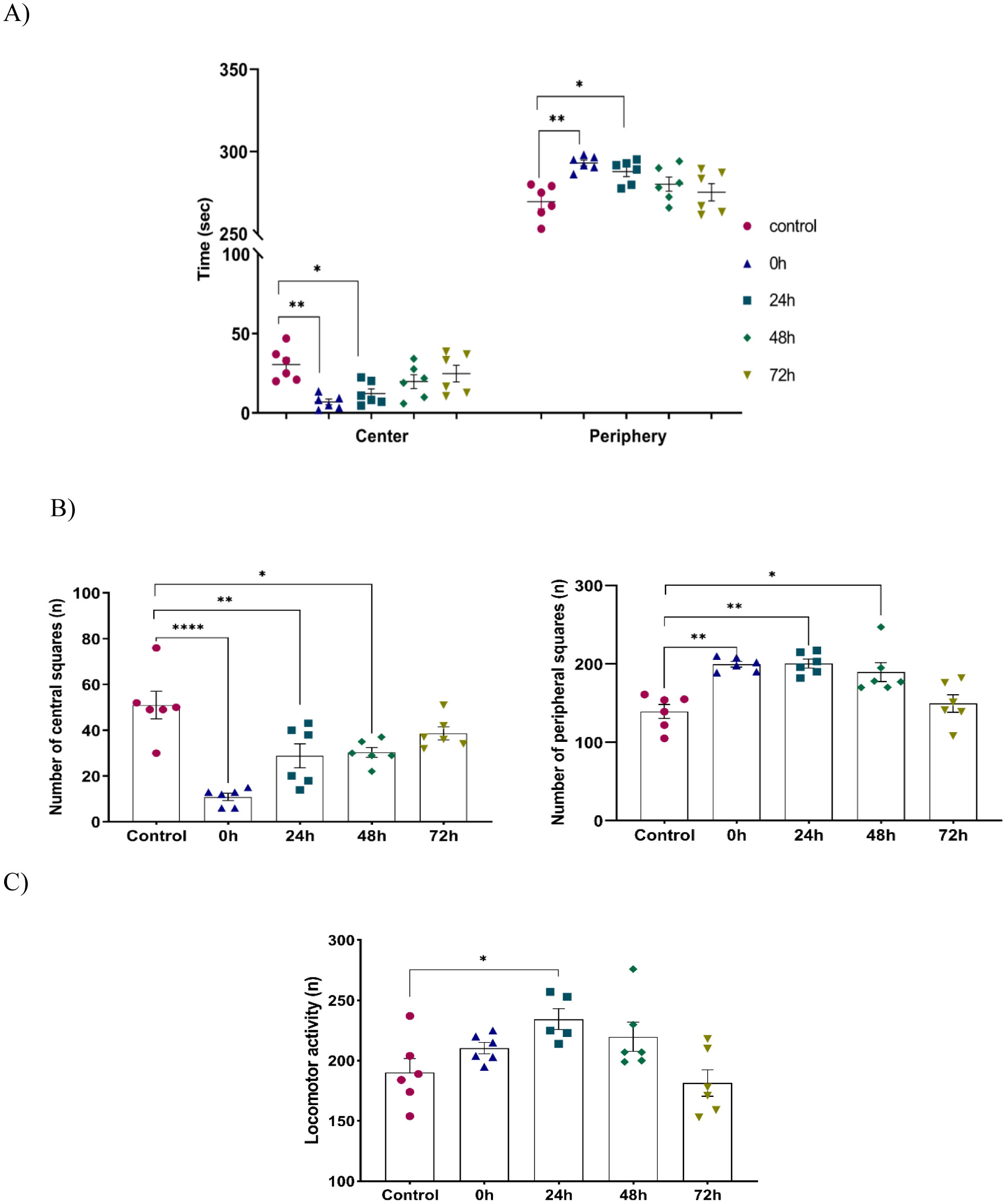
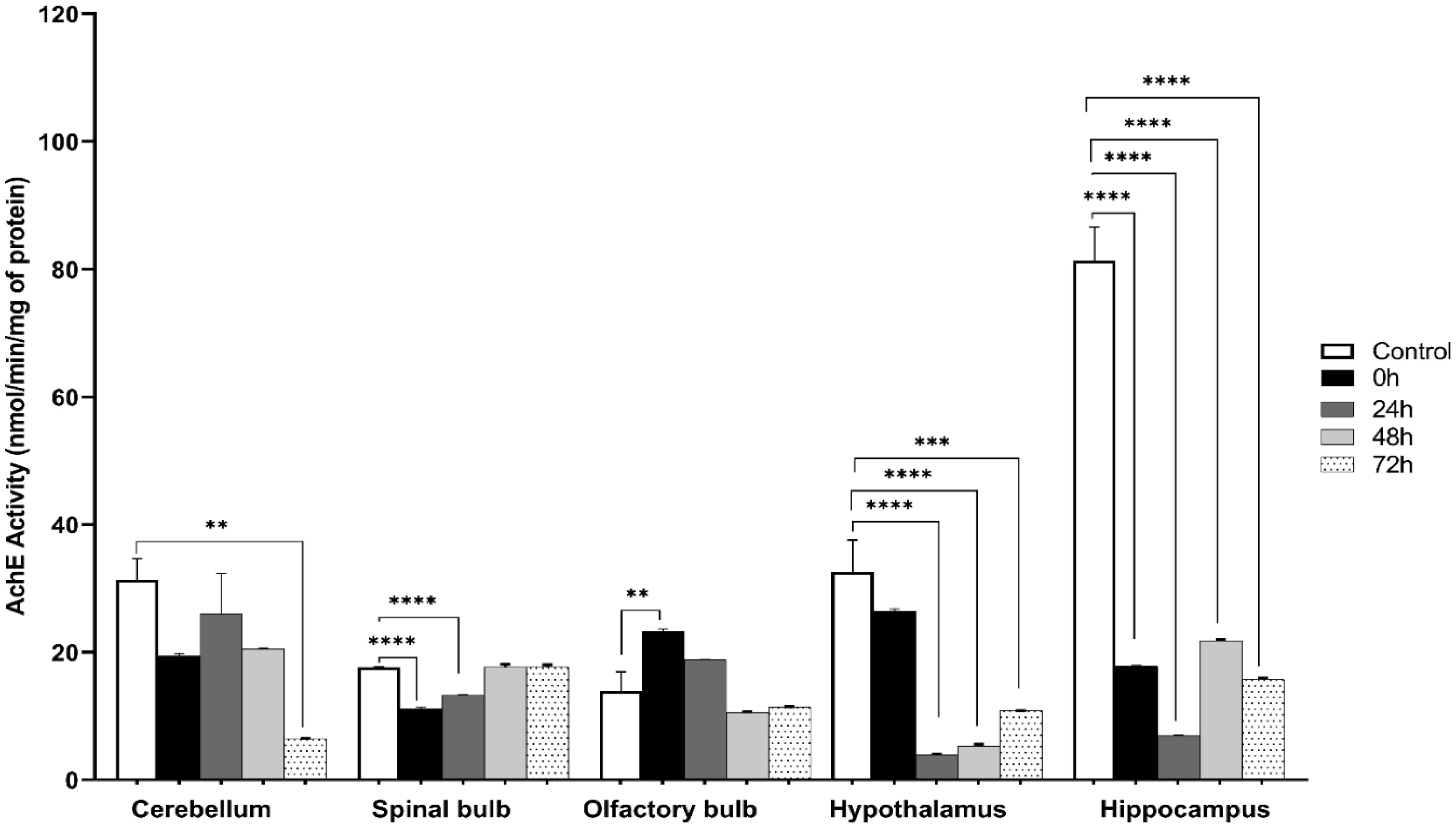
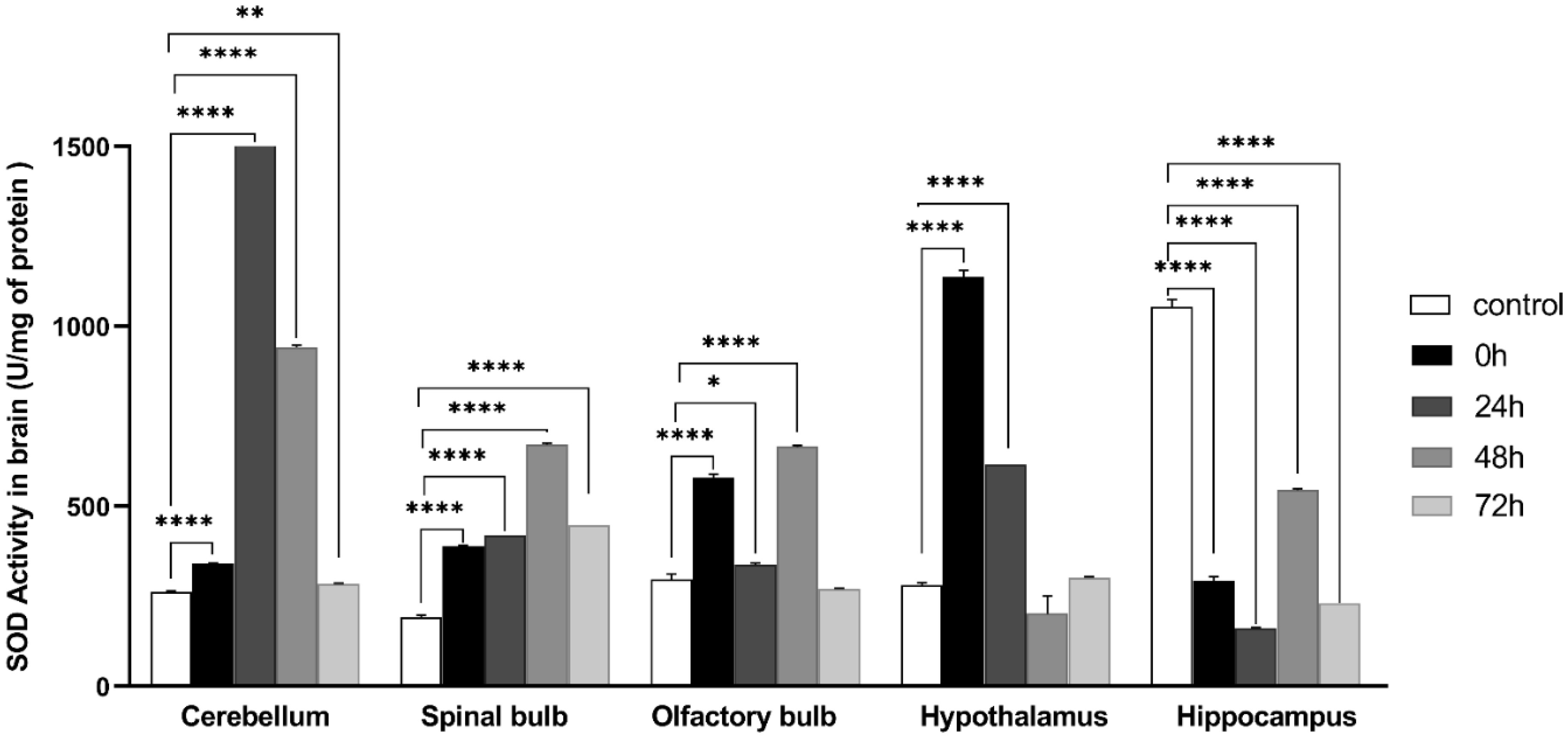
Exposure to certain acute stressors results in an immediate behavioral and physiological response to these situations during a significant period of days. The goal of the current study is to evaluate the long-lasting effect of single exposure of restraint stress among mice after 0 h, 24 h, 48 h and 72 h. Five groups of mice are under experiment: a control group and four groups exposed to one session of restraint stress. All these groups have been studied for behavioral tests in order to evaluate their memories. This is done through a Y-labyrinth and an object recognition test, and anxiety by using open field device. In the second part of the study, enzymatic assays (concerning catalase, glutathione s transferase, glutathione peroxidase and superoxide dismutase) are used to evaluate oxidative stress. The enzymatic activity of the antioxidant system is assessed in five brain structures, including the cerebellum, olfactory bulb, spinal bulb, hypothalamus, and hippocampus.
The obtained results show that acute restraint stress leads to a decrease in memory function and to the development of an anxious state; concomitant to an increase of locomotor activity afterword. It causes disturbance of antioxidant balance in the brain by developing a state of oxidative stress. Indeed, restraint stress causes a change in anti-oxidant stress enzymatic activity in the brain, notably in post-stress period. In conclusion, acute restraint stress is responsible for altering cognitive functions, especially memory, and the development of anxious behavior, which could be a result of the generation of oxidative stress; effects that are persistent over an important period after the cessation of stress.
Citation: Nouhaila Chaoui, Hammou Anarghou, Meriem Laaroussi, Oumaima Essaidi, Mohamed Najimi, Fatiha Chigr. Long lasting effect of acute restraint stress on behavior and brain anti-oxidative status[J]. AIMS Neuroscience, 2022, 9(1): 57-75. doi: 10.3934/Neuroscience.2022005
Exposure to certain acute stressors results in an immediate behavioral and physiological response to these situations during a significant period of days. The goal of the current study is to evaluate the long-lasting effect of single exposure of restraint stress among mice after 0 h, 24 h, 48 h and 72 h. Five groups of mice are under experiment: a control group and four groups exposed to one session of restraint stress. All these groups have been studied for behavioral tests in order to evaluate their memories. This is done through a Y-labyrinth and an object recognition test, and anxiety by using open field device. In the second part of the study, enzymatic assays (concerning catalase, glutathione s transferase, glutathione peroxidase and superoxide dismutase) are used to evaluate oxidative stress. The enzymatic activity of the antioxidant system is assessed in five brain structures, including the cerebellum, olfactory bulb, spinal bulb, hypothalamus, and hippocampus.
The obtained results show that acute restraint stress leads to a decrease in memory function and to the development of an anxious state; concomitant to an increase of locomotor activity afterword. It causes disturbance of antioxidant balance in the brain by developing a state of oxidative stress. Indeed, restraint stress causes a change in anti-oxidant stress enzymatic activity in the brain, notably in post-stress period. In conclusion, acute restraint stress is responsible for altering cognitive functions, especially memory, and the development of anxious behavior, which could be a result of the generation of oxidative stress; effects that are persistent over an important period after the cessation of stress.

| [1] |
Lloyd C, Smith J, Weinger K (2005) Stress and Diabetes: A Review of the Links. Diabetes Spectrum 18: 121-127. https://doi.org/10.2337/diaspect.18.2.121 
|
| [2] |
Manni L, Fausto VD, Fiore M, et al. (2008) Repeated Restraint and Nerve Growth Factor Administration in Male and Female Mice: Effect on Sympathetic and Cardiovascular Mediators of the Stress Response. Curr Neurovasc Res 5: 1-12. https://doi.org/10.2174/156720208783565654
|
| [3] |
Wirtz PH, Redwine LS, Baertschi C, et al. (2008) Coagulation activity before and after acute psychosocial stress increases with age. Psychosom Med 70: 476-481. https://doi.org/10.1097/PSY.0b013e31816e03a5
|
| [4] |
Quick SK, Shields PG, Nie J, et al. (2008) Effect modification by catalase genotype suggests a role for oxidative stress in the association of hormone replacement therapy with postmenopausal breast cancer risk. Cancer Epidemiol Biomarkers Prev 17: 1082-1087. https://doi.org/10.1158/1055-9965.EPI-07-2755
|
| [5] |
Chaplin TM, Hong K, Bergquist K, et al. (2008) Gender differences in response to emotional stress: an assessment across subjective, behavioral, and physiological domains and relations to alcohol craving. Alcohol Clin Exp Res 32: 1242-1250. https://doi.org/10.1111/j.1530-0277.2008.00679.x
|
| [6] |
Harris RBS, Mitchell TD, Simpson J, et al. (2002) Weight loss in rats exposed to repeated acute restraint stress is independent of energy or leptin status. Am J Physiol Regulatory Integrative Comp Physiol 282: R77-R88. https://doi.org/10.1152/ajpregu.2002.282.1.R77
|
| [7] |
Hamer M, Stamatakis E (2008) Inflammation as an intermediate pathway in the association between psychosocial stress and obesity. Physiol Behav 94: 536-539. https://doi.org/10.1016/j.physbeh.2008.03.010
|
| [8] |
Vallès A, Martí O, García A, et al. (2000) Single exposure to stressors causes long-lasting, stress-dependent reduction of food intake in rats. Am J Physiol Regulatory Integrative Comp Physiol 279: R1138-R1144. https://doi.org/10.1152/ajpregu.2000.279.3.R1138
|
| [9] |
Charrier C, Chigr F, Tardivel C, et al. (2006) BDNF regulation in the rat dorsal vagal complex during stress-induced anorexia. Brain Res 1107: 52-57. https://doi.org/10.1016/j.brainres.2006.05.099
|
| [10] |
Adam TC, Epel ES (2007) Stress, eating and the reward system. Physiol Behav 91: 449-458. https://doi.org/10.1016/j.physbeh.2007.04.011
|
| [11] |
Laurent L, Jean A, Manrique C (2013) Anorexia and drugs of abuse abnormally suppress appetite, the result of a shared molecular signal foul-up. Animal Models of Eating Disorders . Totowa: Humana Press 319-331. https://doi.org/10.1007/978-1-62703-104-2_19
|
| [12] |
Gu H, Tang C, Yang Y (2012) Psychological stress, immune response, and atherosclerosis. Atheroscler 223: 69-77. https://doi.org/10.1016/j.atherosclerosis.2012.01.021
|
| [13] |
Paskitti ME, McCreary BJ, Herman JP (2000) Stress regulation of adrenocorticosteroid receptor gene transcription and mRNA expression in rat hippocampus: time-course analysis. Mol Brain Res 80: 142-152. https://doi.org/10.1016/S0169-328X(00)00121-2
|
| [14] |
Johnson JD, Barnard DF, Kulp AC, et al. (2019) Neuroendocrine regulation of brain cytokines after psychological stress. J Endocr Soc 3: 1302-1320. https://doi.org/10.1210/js.2019-00053
|
| [15] |
Chigr F, Rachidi F, Tardivel C, et al. (2014) Modulation of orexigenic and anorexigenic peptides gene expression in the rat DVC and hypothalamus by acute immobilization stress. Front Cell Neurosci 8: 198. https://doi.org/10.3389/fncel.2014.00198
|
| [16] |
Hayashi T (2014) Conversion of psychological stress into cellular stress response: Roles of the sigma-1 receptor in the process. Psychiatry Clin Neurosci 69: 179-191. https://doi.org/10.1111/pcn.12262
|
| [17] |
Chaudhari N, Talwar P, Parimisetty A, et al. (2014) A molecular web: endoplasmic reticulum stress, inflammation, and oxidative stress. Front Cell Neurosci 8: 213. https://doi.org/10.3389/fncel.2014.00213
|
| [18] |
Santos CXC, Tanaka LY, Wosniak JJ, et al. (2009) Mechanisms and implications of reactive oxygen species generation during the unfolded protein response: roles of endoplasmic reticulum oxidoreductases, mitochondrial electron transport, and NADPH oxidase. Antioxid Redox Signal 11: 2409-2427. https://doi.org/10.1089/ars.2009.2625
|
| [19] |
Bernasconi R, Molinari M (2011) ERAD and ERAD tuning: disposal of cargo and of ERAD regulators from the mammalian ER. Curr Opin Cell Biol 23: 176-183. https://doi.org/10.1016/j.ceb.2010.10.002
|
| [20] |
Chaudhuri O, Koshy ST, Da Cunha CB, et al. (2014) Extracellular matrix stiffness and composition jointly regulate the induction of malignant phenotypes in mammary epithelium. Nat Mater 13: 970-8. https://doi.org/10.1038/NMAT4009
|
| [21] |
Ellman GL, Courtney KD, Andres JV, et al. (1961) A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem Pharmacol 7: 88-95. https://doi.org/10.1016/0006-2952(61)90145-9
|
| [22] |
Buege JA, Aust SD (1978) Microsomal lipid peroxidation. Methods Enzymol 105: 302-310. https://doi.org/10.1016/S0076-6879(78)52032-6
|
| [23] |
Aebi H (1974) Catalase. Methods Enzym Anal 2: 673-684. https://doi.org/10.1016/B978-0-12-091302-2.50032-3
|
| [24] |
Flohe L, Gunzler WA (1984) Assays of glutathione peroxidase. Methods Enzymol 105: 114-121. https://doi.org/10.1016/S0076-6879(84)05015-1
|
| [25] |
Habig WH, Pabst MJ, Jakoby WB (1974) Glutathione-S-transferase the first step in mercapturic acid formation. J Biol Chem 249: 7130-9.
|
| [26] |
Asada K, Takahashi M, Nagate M (1974) Assay and inhibitors of spinach superoxide dismutase. Agric Biol Chem 38: 471-473. https://doiorg/10.1080/00021369.1974.10861178
|
| [27] |
Lowry OH, Rosebrough NJ, Farr AL, et al. (1951) Protein measurement with the Folin phenol reagent. J Biol Chem 193: 265-275.
|
| [28] |
Kovács LÁ, Schiessl JA, Nafz AE, et al. (2018) Both basal and acute restraint stress-induced c-Fos expression is influenced by age in the extended amygdala and brainstem stress centers in male rats. Front Aging Neurosci 10: 248. https://doi.org/10.3389/fnagi.2018.00248
|
| [29] |
Bland ST, Schmid MJ, Der-Avakian A, et al. (2005) Expression of c-fos and BDNF mRNA in subregions of the prefrontal cortex of male and female rats after acute uncontrollable stress. Brain Res 1051: 90-99. https://doi.org/10.1016/j.brainres.2005.05.065
|
| [30] | Migdal C, Serres M (2011) Espèces réactives de l'oxygène et stress oxydant. Med Sci 27: 405-412. https://doi.org/10.1051/medsci/2011274017 |
| [31] |
Cherian DA, Peter T, Narayanan A, et al. (2019) Malondialdehyde as a marker of oxidative stress in periodontitis patients. J Pharm Bioallied Sci 11: S297-S300. https://doi.org/10.4103/JPBS.JPBS_17_19
|
| [32] | Tirani MM, Haghjou MM (2019) Reactive oxygen species (ROS), total antioxidant capacity (AOC) and malondialdehyde (MDA) make a triangle in evaluation of zinc stress extension. J Anim Plant Sci 29: 1100-1111. |
| [33] |
Čolović M, Krstić D, Petrović S, et al. (2010) Toxic effects of diazinon and its photodegradation products. Toxicol Lett 193: 9-18. https://doi.org/10.1016/j.toxlet.2009.11.022
|
| [34] |
Akhgari M, Abdollahi M, Kebryaeezadeh A, et al. (2003) Biochemical evidence for free radicalinduced lipid peroxidation as a mechanism for subchronic toxicity of malathion in blood and liver of rats. Hum Exp Toxicol 22: 205-11. https://doi.org/10.1191/0960327103ht346oa
|
| [35] | Abdollahi M, Ranjbar A, Shadnia S, et al. (2004) Pesticides and oxidative stress: a review. Med Sci Monit 10: 141-7. |
| [36] |
Dal-Zotto S, Martí O, Delgado R, et al. (2004) Potentiation of glucocorticoid release does not modify the long-term effects of a single exposure to immobilization stress. Psychopharmacology 177: 230-237. https://doi.org/10.1007/s00213-004-1939-y
|
| [37] |
Akkerman S, Blokland A, Reneerkens O, et al. (2012) Object recognition testing: Methodological considerations on exploration and discrimination measures. Behav Brain Res 232: 335-347. https://doi.org/10.1016/j.bbr.2012.03.022
|
| [38] |
Parent MB, Baxter MG (2004) Septohippocampal Acetylcholine: Involved in but not Necessary for Learning and Memory?. Learn Mem 11: 9-20. https://doi.org/10.1101/lm.69104
|
| [39] |
Micheau J, Marighetto A (2011) Acetylcholine and memory: A long, complex and chaotic but still living relationship. Behav Brain Res 221: 424-429. https://doi.org/10.1016/j.bbr.2010.11.052
|
| [40] |
Deschamps R, Moulignier A (2005) La mémoire et ses troubles Memory and related disorders. EMC- Neurol 2: 505-525. https://doi.org/10.1016/j.emcn.2005.07.003
|
| [41] |
Hasselmo ME (2006) The role of acetylcholine in learning and memory. Curr Opin Neurobiol 6: 710-715. https://doi.org/10.1016/j.conb.2006.09.002
|
| [42] |
Picciotto MR, Higley MJ, Mineur YS (2012) Acetylcholine as a neuromodulator: cholinergic signaling shapes nervous system function and behavior. Neuron 76: 116-129. https://doi.org/10.1016/j.neuron.2012.08.036
|
| [43] |
Haam J, Yakel JL (2017) Cholinergic modulation of the hippocampal region and memory function. J Neurochem 142: 111-121. https://doi.org/10.1111/jnc.14052
|
| [44] |
Newman EL, Gupta K, Climer JR, et al. (2012) Cholinergic modulation of cognitive processing: insights drawn from computational models. Front Behav Neurosci 6: 24. https://doi.org/10.3389/fnbeh.2012.00024
|
| [45] |
Torres LF, Duchen LW (1987) The mutant mdx: inherited myopathy in the mouse: morphological studies of nerves, muscles and end-plates. Brain 110: 269-99. https://doi.org/10.1093/brain/110.2.269
|
| [46] |
Hedner M, Larsson M, Arnold N, et al. (2010) Cognitive factors in odor detection, odor discrimination, and odor identification tasks. J Clin Exp Neuropsychol 32: 1062-7. https://doi.org/10.1080/13803391003683070
|
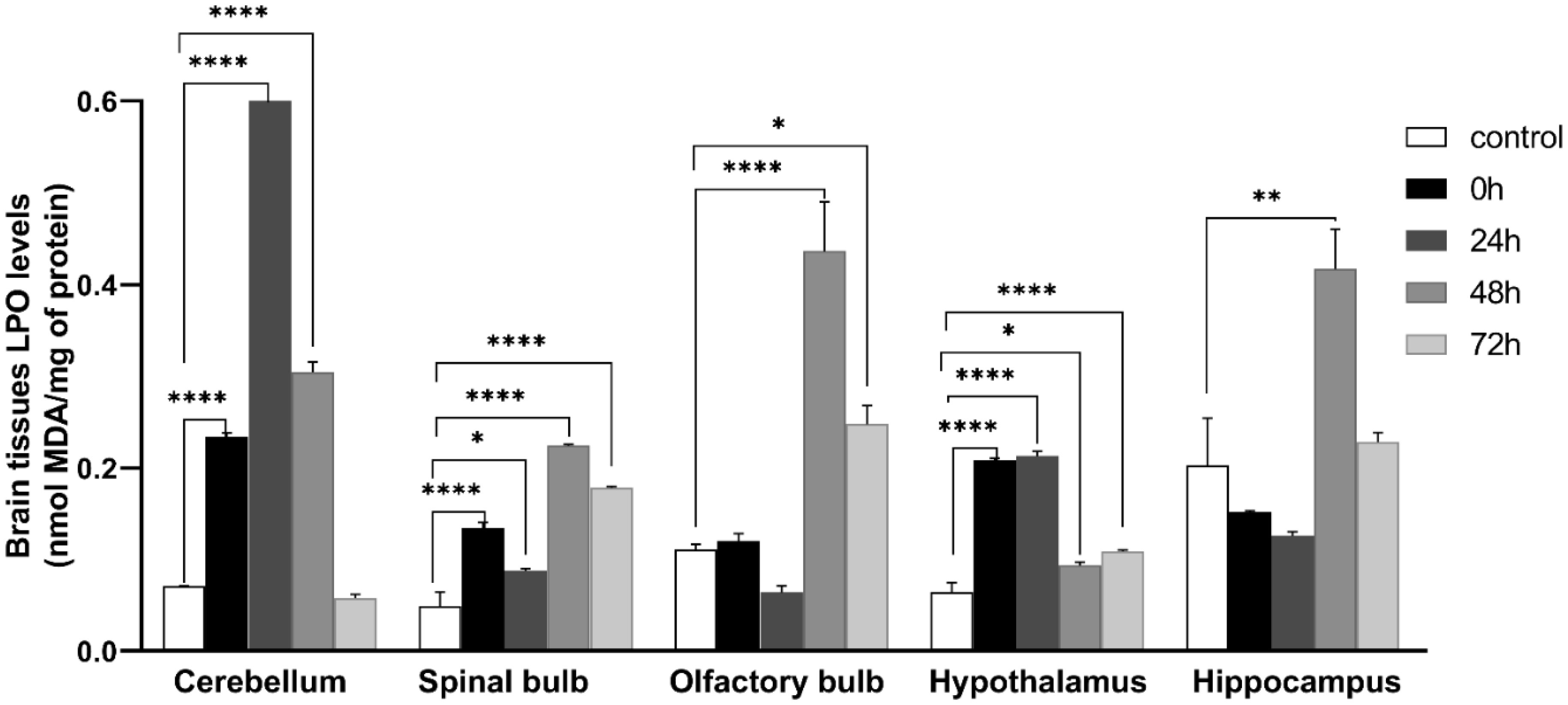
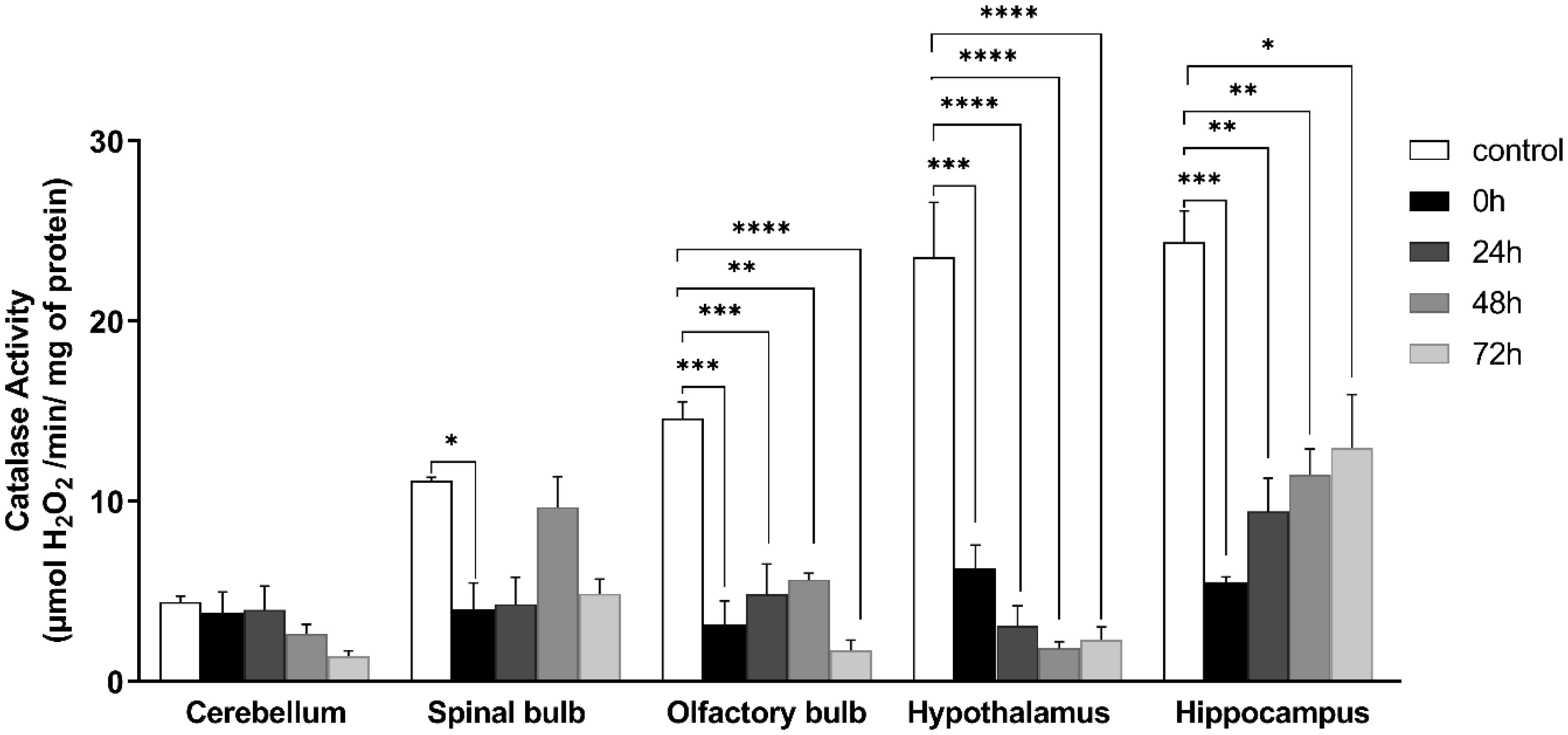
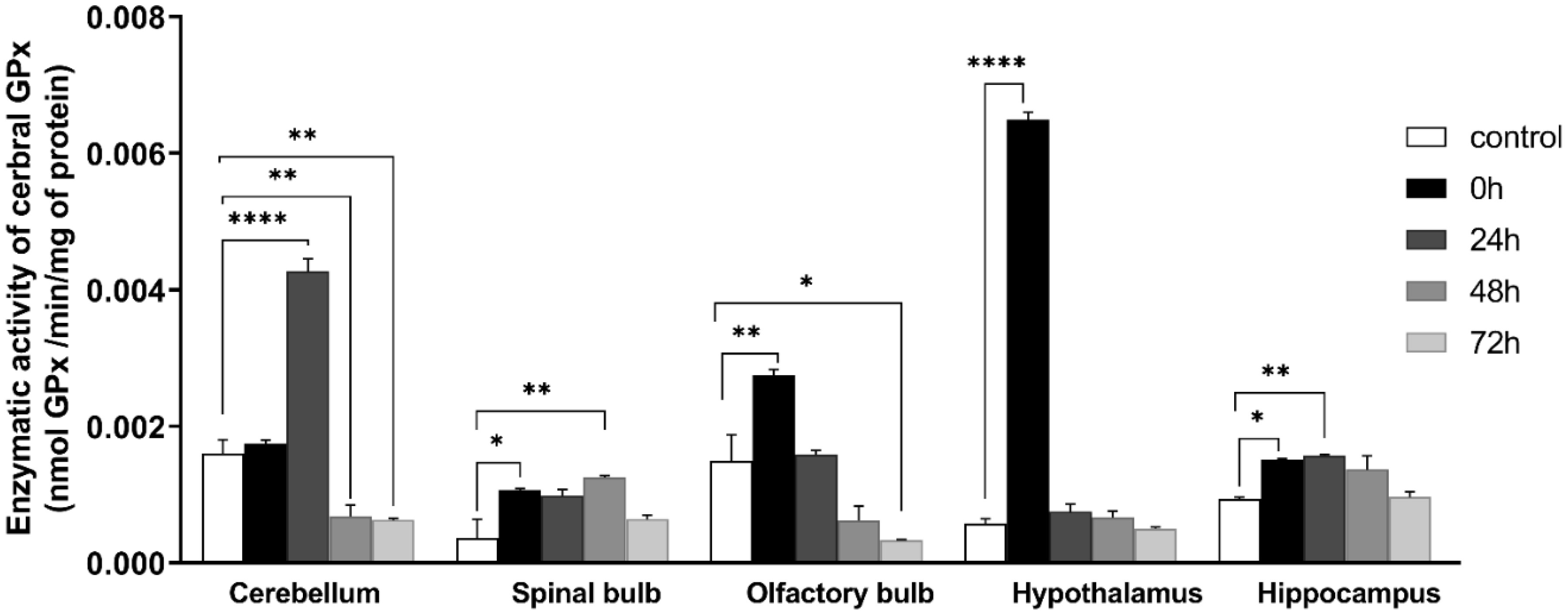
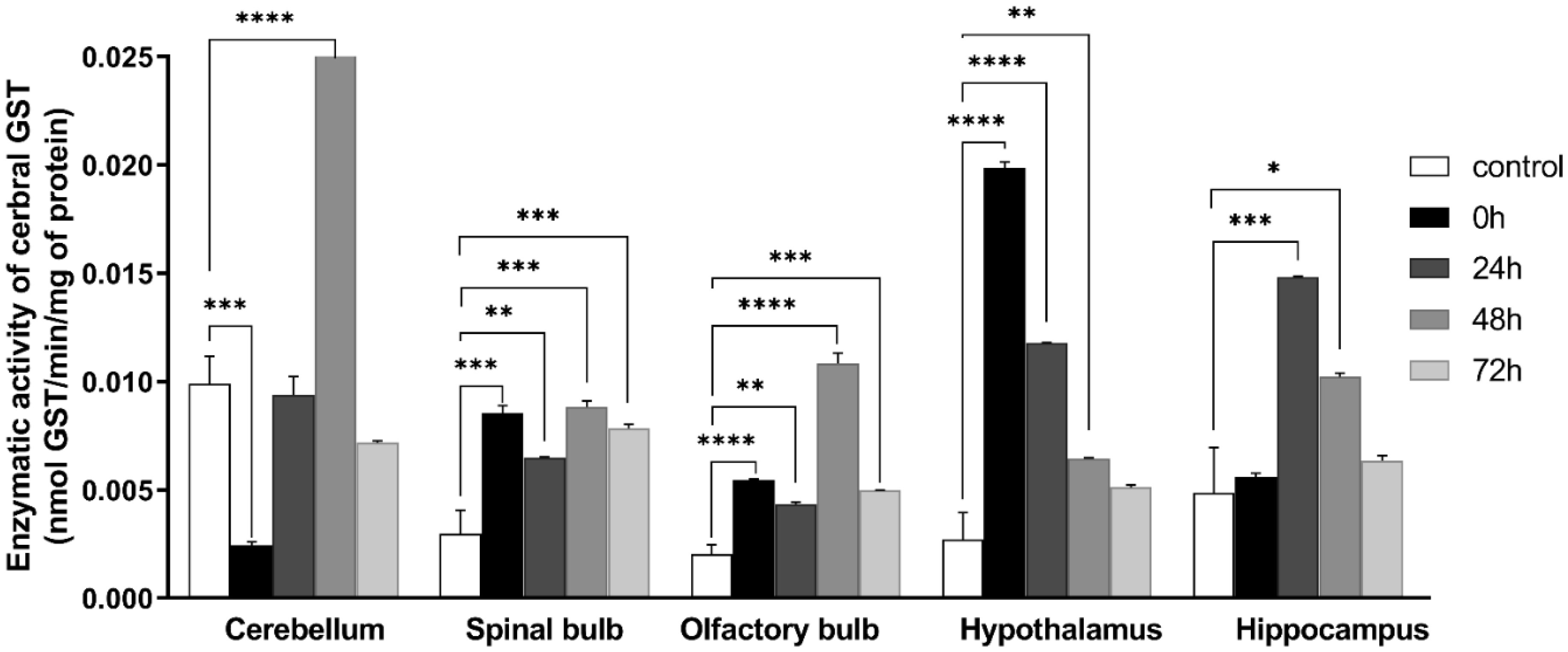
Figures(9)

Nouhaila Chaoui, Hammou Anarghou, Meriem Laaroussi, Oumaima Essaidi, Mohamed Najimi, Fatiha Chigr. Long lasting effect of acute restraint stress on behavior and brain anti-oxidative status[J]. AIMS Neuroscience, 2022, 9(1): 57-75. doi: 10.3934/Neuroscience.2022005







 DownLoad:
DownLoad: