Citation: Dirk Roosterman, Graeme S. Cottrell. Astrocytes and neurons communicate via a monocarboxylic acid shuttle[J]. AIMS Neuroscience, 2020, 7(2): 94-106. doi: 10.3934/Neuroscience.2020007

| [1] |
Pellerin L, Magistretti PJ (2003) Food for thought: challenging the dogmas. J Cereb Blood Flow Metab 23: 1282-1286. doi: 10.1097/01.WCB.0000096064.12129.3D

|
| [2] |
Pellerin L, Magistretti PJ (1994) Glutamate uptake into astrocytes stimulates aerobic glycolysis: a mechanism coupling neuronal activity to glucose utilization. Proc Natl Acad Sci U S A 91: 10625-10629. doi: 10.1073/pnas.91.22.10625
|
| [3] |
Simpson IA, Carruthers A, Vannucci SJ (2007) Supply and demand in cerebral energy metabolism: the role of nutrient transporters. J Cereb Blood Flow Metab 27: 1766-1791. doi: 10.1038/sj.jcbfm.9600521
|
| [4] |
Prigogine I (1978) Time, structure, and fluctuations. Science (New York, NY) 201: 777-785. doi: 10.1126/science.201.4358.777
|
| [5] | Morowitz HJ (1992) Beginnings of Cellular Life: Metabolism Recapitulates Biogenesis Yale University Press. |
| [6] |
Jørgensen SE (1999) Tentative fourth law of thermodynamics applied to description of ecosystem development. Ann N Y Acad Sci 879: 320-343. doi: 10.1111/j.1749-6632.1999.tb10438.x
|
| [7] |
Roosterman D, Meyerhof W, Cottrell GS (2018) Proton transport chains in glucose metabolism: mind the proton. Front Neurosci 12: 404. doi: 10.3389/fnins.2018.00404
|
| [8] |
Crane RK, Krane SM (1956) On the mechanism of the intestinal absorption of sugars. Biochim Biophys Acta 20: 568-569. doi: 10.1016/0006-3002(56)90361-4
|
| [9] |
Hamilton KL (2013) Robert K. Crane—Na+-glucose cotransporter to cure? Front Physiol 4: 53. doi: 10.3389/fphys.2013.00053
|
| [10] | Hebb DO (1949) The Organization of Behavior: A Neuropsychological Theory Wiley: 368. |
| [11] |
Ho M-W, Ulanowicz R (2005) Sustainable systems as organisms? Biosystems 82: 39-51. doi: 10.1016/j.biosystems.2005.05.009
|
| [12] |
Bonen A (2001) The expression of lactate transporters (MCT1 and MCT4) in heart and muscle. Eur J Appl Physiol 86: 6-11. doi: 10.1007/s004210100516
|
| [13] |
Halestrap AP (2013) Monocarboxylic acid transport. Compr Physiol 3: 1611-1643. doi: 10.1002/cphy.c130008
|
| [14] |
Van Hee VF, Labar D, Dehon G, et al. (2017) Radiosynthesis and validation of (+/−)-[18F]-3-fluoro-2-hydroxypropionate ([18F]-FLac) as a PET tracer of lactate to monitor MCT1-dependent lactate uptake in tumors. Oncotarget 8: 24415-24428. doi: 10.18632/oncotarget.14705
|
| [15] |
Pullen TJ, Sylow L, Sun G, et al. (2012) Overexpression of monocarboxylate transporter-1 (SLC16A1) in mouse pancreatic β-cells leads to relative hyperinsulinism during exercise. Diabetes 61: 1719-1725. doi: 10.2337/db11-1531
|
| [16] |
Becker HM, Klier M, Schüler C, et al. (2011) Intramolecular proton shuttle supports not only catalytic but also noncatalytic function of carbonic anhydrase II. Proc Natl Acad Sci U S A 108: 3071-3076. doi: 10.1073/pnas.1014293108
|
| [17] |
Noor SI, Dietz S, Heidtmann H, et al. (2015) Analysis of the binding moiety mediating the interaction between monocarboxylate transporters and carbonic anhydrase II. J Biol Chem 290: 4476-4486. doi: 10.1074/jbc.M114.624577
|
| [18] |
Noor SI, Jamali S, Ames S, et al. (2018) A surface proton antenna in carbonic anhydrase II supports lactate transport in cancer cells. Elife 7: e35176. doi: 10.7554/eLife.35176
|
| [19] |
Dimmer KS, Friedrich B, Lang F, et al. (2000) The low-affinity monocarboxylate transporter MCT4 is adapted to the export of lactate in highly glycolytic cells. Biochem J 350: 219-227. doi: 10.1042/bj3500219
|
| [20] |
Lynch CJ, Xu Y, Hajnal A, et al. (2015) RNA sequencing reveals a slow to fast muscle fiber type transition after olanzapine infusion in rats. PloS One 10: e0123966. doi: 10.1371/journal.pone.0123966
|
| [21] |
Pierre K, Magistretti PJ, Pellerin L (2002) MCT2 is a major neuronal monocarboxylate transporter in the adult mouse brain. J Cereb Blood Flow Metab 22: 586-595. doi: 10.1097/00004647-200205000-00010
|
| [22] |
Forero-Quintero LS, Ames S, Schneider H-P, et al. (2019) Membrane-anchored carbonic anhydrase IV interacts with monocarboxylate transporters via their chaperones CD147 and GP70. J Biol Chem 294: 593-607. doi: 10.1074/jbc.RA118.005536
|
| [23] |
Klier M, Schüler C, Halestrap AP, et al. (2011) Transport activity of the high-affinity monocarboxylate transporter MCT2 is enhanced by extracellular carbonic anhydrase IV but not by intracellular carbonic anhydrase II. J Biol Chem 286: 27781-27791. doi: 10.1074/jbc.M111.255331
|
| [24] |
Itel F, Al-Samir S, Öberg F, et al. (2012) CO2 permeability of cell membranes is regulated by membrane cholesterol and protein gas channels. FASEB J 26: 5182-5191. doi: 10.1096/fj.12-209916
|
| [25] |
Cornelius F (2001) Modulation of Na,K-ATPase and Na-ATPase activity by phospholipids and cholesterol. I. Steady-state kinetics. Biochemistry 40: 8842-8851. doi: 10.1021/bi010541g
|
| [26] |
Broer S, Broer A, Schneider HP, et al. (1999) Characterization of the high-affinity monocarboxylate transporter MCT2 in Xenopus laevis oocytes. Biochem J 341: 529-535. doi: 10.1042/bj3410529
|
| [27] |
Lin RY, Vera JC, Chaganti RS, et al. (1998) Human monocarboxylate transporter 2 (MCT2) is a high affinity pyruvate transporter. J Biol Chem 273: 28959-28965. doi: 10.1074/jbc.273.44.28959
|
| [28] | Gibbs ME (2015) Role of glycogenolysis in memory and learning: regulation by noradrenaline, serotonin and ATP. Front Integr Neurosci 9: 70. |
| [29] |
Magistretti PJ, Allaman I (2015) A cellular perspective on brain energy metabolism and functional imaging. Neuron 86: 883-901. doi: 10.1016/j.neuron.2015.03.035
|
| [30] |
Newman LA, Korol DL, Gold PE (2011) Lactate produced by glycogenolysis in astrocytes regulates memory processing. PloS One 6: e28427. doi: 10.1371/journal.pone.0028427
|
| [31] |
Suzuki A, Stern SA, Bozdagi O, et al. (2011) Astrocyte-neuron lactate transport is required for long-term memory formation. Cell 144: 810-823. doi: 10.1016/j.cell.2011.02.018
|
| [32] |
Bröer S, Schneider HP, Bröer A, et al. (1998) Characterization of the monocarboxylate transporter 1 expressed in Xenopus laevis oocytes by changes in cytosolic pH. Biochem J 333: 167-174. doi: 10.1042/bj3330167
|
| [33] |
de Bruijne AW, Vreeburg H, van Steveninck J (1985) Alternative-substrate inhibition of L-lactate transport via the monocarboxylate-specific carrier system in human erythrocytes. Biochim Biophys Acta 812: 841-844. doi: 10.1016/0005-2736(85)90280-9
|
| [34] |
Sahlin K, Harris RC, Nylind B, et al. (1976) Lactate content and pH in muscle obtained after dynamic exercise. Pflugers Arch 367: 143-149. doi: 10.1007/BF00585150
|
| [35] |
Urbańska K, Orzechowski A (2019) Unappreciated role of LDHA and LDHB to control apoptosis and autophagy in tumor cells. Int J Mol Sci 20: E2085. doi: 10.3390/ijms20092085
|
| [36] | Svedružić ŽM, Odorčić I, Chang CH, et al. (2020) Substrate channeling via a transient protein-protein complex: the case of D-Glyceraldehyde-3-Phosphate dehydrogenase and L-Lactate dehydrogenase. bioRxiv 2020.2001.2022.916023. |
| [37] |
Svedruzić ZM, Spivey HO (2006) Interaction between mammalian glyceraldehyde-3-phosphate dehydrogenase and L-lactate dehydrogenase from heart and muscle. Proteins 63: 501-511. doi: 10.1002/prot.20862
|
| [38] |
Ovádi J, Srere PA (1992) Channel your energies. Trends Biochem Sci 17: 445-447. doi: 10.1016/0968-0004(92)90485-R
|
| [39] |
Reed CA (2013) Myths about the proton. The nature of H+ in condensed media. Acc Chem Res 46: 2567-2575. doi: 10.1021/ar400064q
|
| [40] |
Hashimoto T, Hussien R, Brooks GA (2006) Colocalization of MCT1, CD147, and LDH in mitochondrial inner membrane of L6 muscle cells: evidence of a mitochondrial lactate oxidation complex. Am J Physiol Endocrinol Metab 290: E1237-1244. doi: 10.1152/ajpendo.00594.2005
|
| [41] |
Fredriksson S, Gullberg M, Jarvius J, et al. (2002) Protein detection using proximity-dependent DNA ligation assays. Nat Biotechnol 20: 473-477. doi: 10.1038/nbt0502-473
|
| [42] |
Soderberg O, Leuchowius KJ, Gullberg M, et al. (2008) Characterizing proteins and their interactions in cells and tissues using the in situ proximity ligation assay. Methods 45: 227-232. doi: 10.1016/j.ymeth.2008.06.014
|
| [43] |
Brooks GA (2020) Lactate as a fulcrum of metabolism. Redox Biol 101454. doi: 10.1016/j.redox.2020.101454
|
| [44] |
Tadi M, Allaman I, Lengacher S, et al. (2015) Learning-Induced gene expression in the hippocampus reveals a role of neuron -astrocyte metabolic coupling in long term memory. PloS One 10: e0141568. doi: 10.1371/journal.pone.0141568
|
| [45] |
Contreras-Baeza Y, Sandoval PY, Alarcón R, et al. (2019) Monocarboxylate transporter 4 (MCT4) is a high affinity transporter capable of exporting lactate in high-lactate microenvironments. J Biol Chem 294: 20135-20147. doi: 10.1074/jbc.RA119.009093
|
| [46] |
Takanaga H, Chaudhuri B, Frommer WB (2008) GLUT1 and GLUT9 as major contributors to glucose influx in HepG2 cells identified by a high sensitivity intramolecular FRET glucose sensor. Biochim Biophys Acta 1778: 1091-1099. doi: 10.1016/j.bbamem.2007.11.015
|
| [47] |
Nedergaard M, Ransom B, Goldman SA (2003) New roles for astrocytes: redefining the functional architecture of the brain. Trends Neurosci 26: 523-530. doi: 10.1016/j.tins.2003.08.008
|
| [48] |
Korogod N, Petersen CCH, Knott GW (2015) Ultrastructural analysis of adult mouse neocortex comparing aldehyde perfusion with cryo fixation. Elife 4. doi: 10.7554/eLife.05793
|
| [49] | Mason S (2017) Lactate shuttles in neuroenergetics-homeostasis, allostasis and beyond. Front Neurosci 11: 43. |
| [50] |
Pellerin L (2010) Food for thought: the importance of glucose and other energy substrates for sustaining brain function under varying levels of activity. Diabetes Metab 36: S59-63. doi: 10.1016/S1262-3636(10)70469-9
|
| [51] |
Gandhi GK, Cruz NF, Ball KK, et al. (2009) Astrocytes are poised for lactate trafficking and release from activated brain and for supply of glucose to neurons. J Neurochem 111: 522-536. doi: 10.1111/j.1471-4159.2009.06333.x
|
| [52] |
Iliff J, Simon M (2019) CrossTalk proposal: The glymphatic system supports convective exchange of cerebrospinal fluid and brain interstitial fluid that is mediated by perivascular aquaporin-4. J Physiol 597: 4417-4419. doi: 10.1113/JP277635
|
| [53] |
Leen WG, Willemsen MA, Wevers RA, et al. (2012) Cerebrospinal fluid glucose and lactate: age-specific reference values and implications for clinical practice. PloS One 7: e42745. doi: 10.1371/journal.pone.0042745
|
| [54] | Gjedden A, Hansen A, Silver I (1980) The glucose concentration of brain interstitial fluid is low. Proc Int Union Physiol Sci 14. |
| [55] |
Prats C, Graham TE, Shearer J (2018) The dynamic life of the glycogen granule. J Biol Chem 293: 7089-7098. doi: 10.1074/jbc.R117.802843
|
| [56] | Lowry OH, Passonneau JV, Hasselberger FX, et al. (1964) Effect of ischemia on known substrates and cofactors of the glycolytic pathway in brain. J Biol Chem 239: 18-30. |
| [57] |
Scemes E, Giaume C (2006) Astrocyte calcium waves: what they are and what they do. Glia 54: 716-725. doi: 10.1002/glia.20374
|
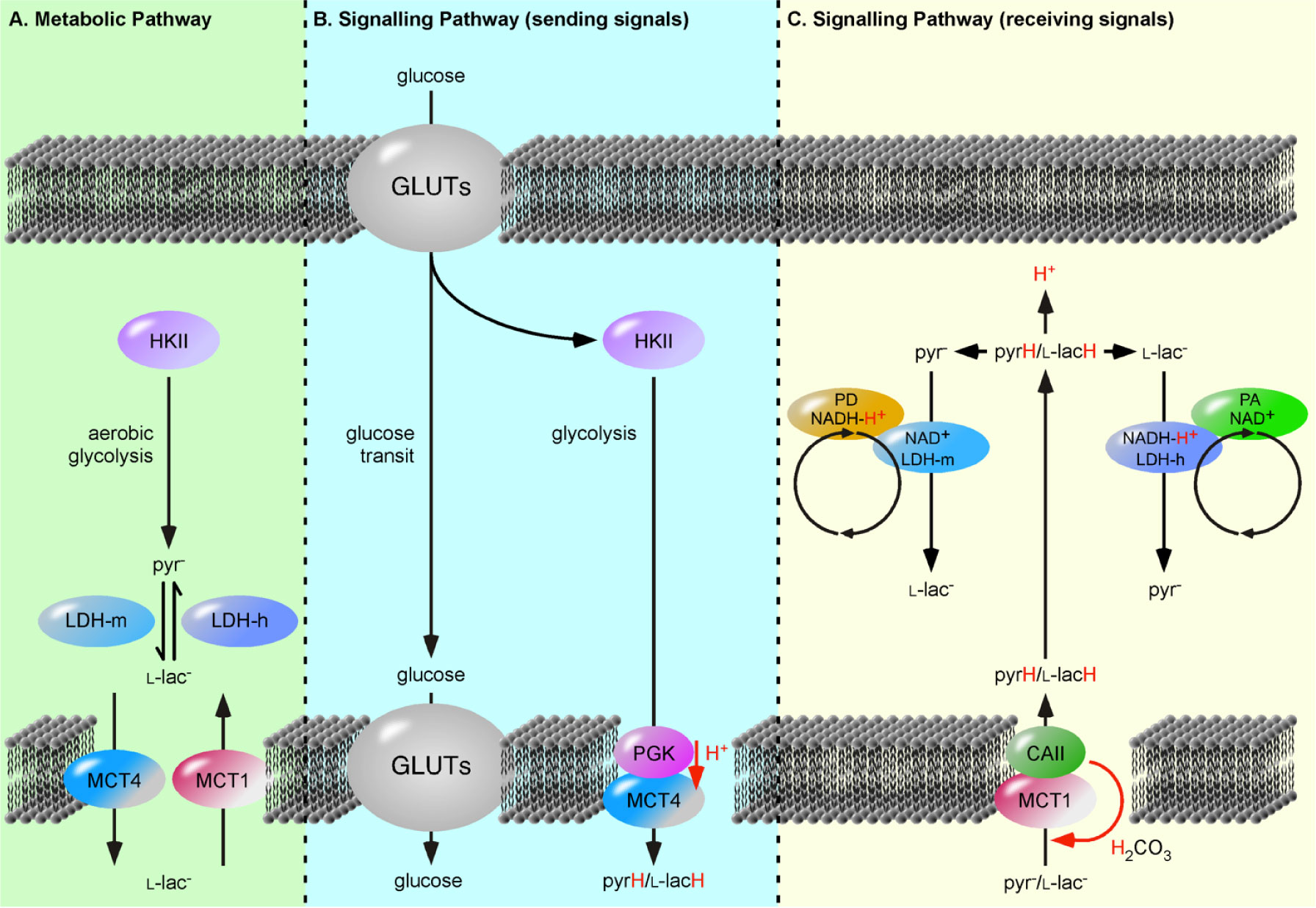
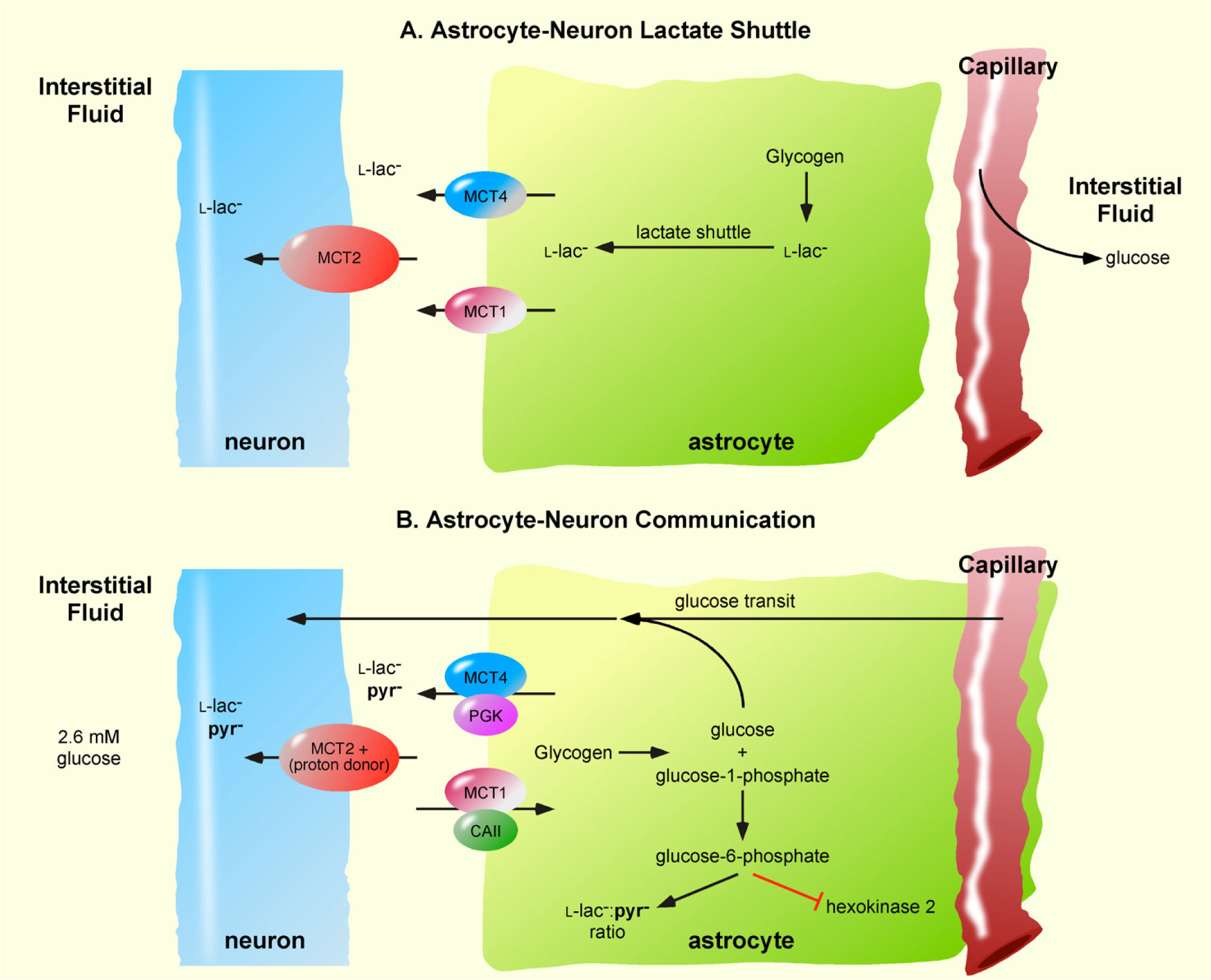
Figures(2) / Tables(1)

Dirk Roosterman, Graeme S. Cottrell. Astrocytes and neurons communicate via a monocarboxylic acid shuttle[J]. AIMS Neuroscience, 2020, 7(2): 94-106. doi: 10.3934/Neuroscience.2020007







 DownLoad:
DownLoad: