Citation: Testimony J Olumade, Oluwafolajimi A Adesanya, Iyanuoluwa J Fred-Akintunwa, David O Babalola, Judith U Oguzie, Olusola A Ogunsanya, Uwem E George, Oluwawapelumi D Akin-Ajani, Damilola G Osasona. Infectious disease outbreak preparedness and response in Nigeria: history, limitations and recommendations for global health policy and practice[J]. AIMS Public Health, 2020, 7(4): 736-757. doi: 10.3934/publichealth.2020057

| [1] | Nigeria Population (2020) Worldometer. Available from: https://www.worldometers.info/world-population/nigeria-population/. |
| [2] |
Muhammad F, Abdulkareem JH, Chowdhury AA (2017) Major Public Health Problems in Nigeria: A review. South East Asia J Public Health 7: 6-11. doi: 10.3329/seajph.v7i1.34672

|
| [3] |
Lecompte E, Fichet-Calvet E, Daffis S, et al. (2006) Mastomys natalensis and Lassa fever, West Africa. Emerg Infect Dis 12: 1971. doi: 10.3201/eid1212.060812
|
| [4] | Emperador DM, Yimer SA, Mazzola LT, et al. (2018) Diagnostic needs for lassa fever outbreak detection, clinical care, and vaccine development. Am J Trop Med Hyg 99: 524-525. |
| [5] |
Mofolorunsho KC (2016) Outbreak of lassa fever in nigeria: Measures for prevention and control. Pan Afr Med J 23. doi: 10.11604/pamj.2016.23.210.8923
|
| [6] | CDC. Lassa Fever Fact Sheet. Available from: https://www.cdc.gov/vhf/lassa/index.html. |
| [7] |
Achinge GI, Kur JT, Gyoh SK (2013) Lassa fever Outbreak in Makurdi, North Central Nigeria: What You Need To Know. IOSR J Dent Med Sci 7: 42-46. doi: 10.9790/0853-0754246
|
| [8] |
Frame JD, Baldwin JM, Gocke DJ, et al. (1970) Lassa fever, a new virus disease of man from West Africa. I. Clinical description and pathological findings. Am J Trop Med Hyg 19: 670-676. doi: 10.4269/ajtmh.1970.19.670
|
| [9] | Ogoina D (2013) Lassa fever: a clinical and epidemiological review. Niger Delta J Med Res 1: 1-10. |
| [10] |
Siddle KJ, Eromon P, Barnes KG, et al. (2018) Genomic Analysis of Lassa Virus during an Increase in Cases in Nigeria in 2018. N Engl J Med 379: 1745-1753. doi: 10.1056/NEJMoa1804498
|
| [11] | Unah L (2020) Nigeria struggles with largest recorded Lassa fever outbreak. Devex. Available from: https://www.devex.com/news/nigeria-struggles-with-largest-recorded-lassa-fever-outbreak-96773. |
| [12] |
Aylward B, Barboza P, Bawo L, et al. (2014) Ebola Virus Disease in West Africa - The First 9 Months of the Epidemic and Forward Projections. N Engl J Med 371: 1481-1495. doi: 10.1056/NEJMoa1411100
|
| [13] |
Folarin OA, Ehichioya D, Schaffner SF, et al. (2016) Ebola Virus Epidemiology and Evolution in Nigeria. J Infect Dis 214: S102-S109. doi: 10.1093/infdis/jiw190
|
| [14] | Otu A, Ameh S, Osifo-Dawodu E, et al. (2017) An account of the Ebola virus disease outbreak in Nigeria: Implications and lessons learnt. BMC Public Health 18: 1-8. |
| [15] | WHO (2014) WHO declares end of Ebola outbreak in Nigeria. Available from: https://www.who.int/mediacentre/news/statements/2014/nigeria-ends-ebola/en/. |
| [16] | Obi R. Yellow Fever Hemorrhagic Disease is just around the corner (1). The Leader Newspaper Online. Available from: https://theleaderassumpta.com/yellow-fever-hemorrhagic-disease-is-just-around-the-corner-1/. |
| [17] | WHO (2015) Increased risk of urban yellow fever outbreaks in Africa. Available from: https://www.who.int/csr/disease/yellowfev/urbanoutbreaks/en/. |
| [18] |
Nwachukwu WE, Yusuff H, Nwangwu U, et al. (2020) The response to re-emergence of yellow fever in Nigeria, 2017. Int J Infect Dis 92: 189-196. doi: 10.1016/j.ijid.2019.12.034
|
| [19] | WHO (2019) Yellow fever - Nigeria. Available from: https://www.who.int/csr/don/09-january-2019-yellow-fever-nigeria/en/. |
| [20] | WHO (2019) Yellow fever - Nigeria. Available from: https://www.who.int/csr/don/17-december-2019-yellow-fever-nigeria/en/. |
| [21] | Nigeria Centre for Disease Control (2020) An update of Yellow Fever outbreak in Nigeria. Available from: https://ncdc.gov.ng/diseases/sitreps/?cat=10&name=An%20update%20of%20Yellow%20Fever%20outbreak%20in%20Nigeria. |
| [22] | Hambrosky J, Kroger A, Wolfe C (2015) Centres for Disease Control and Prevention - “Poliomyelitis”, Epidemiology and Prevention of Vaccine-Preventable Diseases Washington DC: Public Health Foundation. |
| [23] | Last A (2007) Vaccine-linked polio hits Nigeria. BBC News. Available from: http://news.bbc.co.uk/2/hi/africa/7037462.stm. |
| [24] |
Bolu O, Nnadi C, Damisa E, et al. (2018) Progress Toward Poliomyelitis Eradication—Nigeria, January–December 2017. Morb Mortal Wkly Rep 67: 253-256. doi: 10.15585/mmwr.mm6708a5
|
| [25] |
Ogoina D, Izibewule JH, Ogunleye A, et al. (2019) The 2017 human monkeypox outbreak in Nigeria - Report of outbreak experience and response in the Niger Delta University, Bayelsa State, Nigeria. PLoS One 14: e0214229. doi: 10.1371/journal.pone.0214229
|
| [26] | Nigeria Centre for Disease Control. An Update of Monkeypox Outbreak in Nigeria. Available from: https://ncdc.gov.ng/diseases/sitreps/?cat=8&name=An%20Update%20of%20Monkeypox%20Outbreak%20in%20Nigeria. |
| [27] |
Yinka-Ogunleye A, Aruna O, Ogoina D, et al. (2018) Reemergence of human monkeypox in Nigeria, 2017. Emerg Infect Dis 24: 1149. doi: 10.3201/eid2406.180017
|
| [28] | Wilder-Smith A, Leong WY (2017) Importation of yellow fever into China: assessing travel patterns. J Travel Med 24: tax008. |
| [29] |
Njidda AM, Oyebanji O, Obasanya J, et al. (2018) The Nigeria Centre for Disease Control. BMJ Global Health 3: 3-5. doi: 10.1136/bmjgh-2018-000712
|
| [30] | Nigeria Centre for Disease Control. Available from: https://ncdc.gov.ng/. |
| [31] | Nigeria Centre for Disease Control. Field Epidemiology and Laboratory Trainig Program (NFELTP). Available from: https://www.ncdc.gov.ng/training/nfeltp. |
| [32] | Nguku P, Oyemakinde A, Sabitu K, et al. (2014) Training and service in public health, Nigeria Field Epidemiology and Laboratory Training, 2008–2014. Pan Afr Med J 18: 2. |
| [33] | WHO (2014) Nigeria is now free of Ebola virus transmission. Available from: https://www.who.int/mediacentre/news/ebola/20-october-2014/en/. |
| [34] |
Musa E, Nasidi A, Shuaib F, et al. (2016) Nigeria's Ebola outbreak response: lessons for future epidemic preparedness. Arch Med 8. doi: 10.21767/1989-5216.1000171
|
| [35] | Shuaib F, Gunnala R, Musa EO, et al. (2014) Ebola virus disease outbreak—Nigeria, July–September 2014. Morb Mortal Wkly Rep 63: 866-872. |
| [36] | WHO (2015) Ebola situation in Port Harcourt, Nigeria. Available from: https://www.who.int/mediacentre/news/ebola/3-september-2014/en/. |
| [37] | WHO (2015) Ebola in Nigeria and Senegal: stable - for the moment. Available from: https://www.who.int/csr/disease/ebola/ebola-6-months/nigeria-senegal/en/. |
| [38] | Olu OO, Lamunu M, Nanyunja M, et al. (2016) Contact Tracing during an Outbreak of Ebola Virus Disease in the Western Area Districts of Sierra Leone: Lessons for Future Ebola Outbreak Response. Front Public Health 4: 130. |
| [39] | Abdulraheem IS (2002) Public health importance of lassa fever epidemiology, clinical features and current management review of literature. African J Clin Exp Microbiol 3: 33-37. |
| [40] |
Ajayi NA, Nwigwe CG, Azuogu BN, et al. (2013) Containing a Lassa fever epidemic in a resource-limited setting: outbreak description and lessons learned from Abakaliki, Nigeria (January–March 2012). Int J Infect Dis 17: e1011-e1016. doi: 10.1016/j.ijid.2013.05.015
|
| [41] | Dan-nwafor CC, Furuse Y, Ilori EA, et al. (2019) Measures to control protracted large Lassa fever outbreak in Nigeria, 1 January to 28 April 2019. Eurosurveillance 24: 1900272. |
| [42] | Ihekweazu C (2018) National guidelines for lassa fever case management. Abuja. Available from: https://ncdc.gov.ng/themes/common/docs/protocols/92_1547068532.pdf. |
| [43] |
Okokhere P, Colubri A, Azubike C, et al. (2018) Clinical and laboratory predictors of Lassa fever outcome in a dedicated treatment facility in Nigeria: a retrospective, observational cohort study. Lancet Infect Dis 18: 684-695. doi: 10.1016/S1473-3099(18)30121-X
|
| [44] | Monkeypox Outbreak Response: Interim National Guidelines. Abuja; 2017. Available from: https://ncdc.gov.ng/themes/common/docs/protocols/50_1508912430.pdf. |
| [45] |
Ajogbasile FV, Oguzie JU, Oluniyi PE, et al. (2020) Real-time Metagenomic Analysis of Undiagnosed Fever Cases Unveils a Yellow Fever Outbreak in Edo State, Nigeria. Sci Rep 10: 3180. doi: 10.1038/s41598-020-59880-w
|
| [46] | Nigeria's Population Projected to Double by 2050. Voice of America. 2019. Available from: https://www.voanews.com/africa/nigerias-population-projected-double-2050. |
| [47] | Campbell J (2018) Nigeria Faces a Crippling Population Boom. Council on Foreign Relations. 2018. Available from: https://www.cfr.org/blog/nigeria-faces-crippling-population-boom. |
| [48] | Kumar R, Pal R (2018) India achieves WHO recommended doctor population ratio: A call for paradigm shift in public health discourse!. J Fam Med Prim Care 7: 841-844. |
| [49] | World Health Organisation (2006) The World Health Report 2006 - working together for health. Available from: https://www.who.int/whr/2006/en/. |
| [50] | (2011) World Health OrganisationThe Abuja Declaration: Ten Years On. WHO. Geneva: World Health Organization. |
| [51] | PACFAH, CS-SUNN (2017) Budgeting for Health and Nutrition in Nigeria: Trend Analysis. Trend in Health Allocation in Nigeria. Available from: http://www.cs-sunn.org/wp-content/uploads/2017/08/Download-Infographics.pdf. |
| [52] | Adekoya F, Muanya C, Nelson C, et al. (2018) Analysts unveil highs, lows of 2019 budget. Guardian Available from: https://guardian.ng/news/analysts-unveil-highs-lows-of-2019-budget/. |
| [53] | Aworinde T (2019) 2020 budget: Health, education in familiar territory of neglect. Punch Newspapers. Available from: https://punchng.com/2020-budget-health-education-in-familiar-territory-of-neglect/. |
| [54] | Akintomide C, Tapsoba MS, Zhao MF (2014) Strengthening west Africa's public health systems. |
| [55] |
Kirigia JM, Barry SP (2008) Health challenges in Africa and the way forward. Int Arch Med 1: 37-41. doi: 10.1186/1755-7682-1-27
|
| [56] | Uzoechina O (2008) “State Fragility” and the Challenges of Development in West Africa: Moving from Reaction to Prevention. ALC Research Report . |
| [57] | Charrièrre F, Fresia M (2008) West Africa as a Migration and Protection area. UN High Comm Refug. Available from: http://www.refworld.org/docid/4a277db82.html. |
| [58] |
Kakai CG, Okunromade OF, Dan-Nwafor CC, et al. (2020) Improving Cross-Border Preparedness and Response: Lessons Learned from 3 Lassa Fever Outbreaks Across Benin, Nigeria, and Togo, 2017–2019. Health Secur 18: S105-S112. doi: 10.1089/hs.2019.0080
|
| [59] |
Gething PW, Noor AM, Gikandi PW, et al. (2006) Improving imperfect data from health management information systems in Africa using space-time geostatistics. PLoS Med 3: 0825-0831. doi: 10.1371/journal.pmed.0030271
|
| [60] | (2017) World BankFrom Panic and Neglect to Investing in Health Security: Financing Pandemic Preparedness at a National Level. Washington DC: . Report No.: 115271 |
| [61] | Nigeria Center for Disease Control (2018) National Action Plan for Health Security 2018–2022. Abuja. Available from: https://ncdc.gov.ng/themes/common/files/establishment/5e88f9e22d2b4e4563b527005c8a0c43.pdf. |
| [62] |
Talisuna AO, Okiro EA, Yahaya AA, et al. (2020) Spatial and temporal distribution of infectious disease epidemics, disasters and other potential public health emergencies in the World Health Organisation Africa region, 2016–2018. Globalization Health 16: 1-12. doi: 10.1186/s12992-019-0540-4
|
| [63] | Owoseye A (2018) Nigeria needs N134bn to strengthen national health security - Official - Premium Times Nigeria. Available from: https://www.premiumtimesng.com/health/health-news/301626-nigeria-needs-n134bn-to-strengthen-national-health-security-health-official.html. |
| [64] | World Health Organization (2017) Joint External Evaluation of IHR core capacities of the Federal Republic of Nigeria. Geneva. Available from: https://www.who.int/ihr/publications/WHO-WHE-CPI-REP-2017.46/en/. |
| [65] |
Abubakar A, Idris S, Sabitu K, et al. (2010) Emergency preparedness and the capability to identify outbreaks: A case study of Sabon Gari Local Government Area, Kaduna state. Ann Niger Med 4: 21. doi: 10.4103/0331-3131.73877
|
| [66] |
Phalkey RK, Yamamoto S, Awate P, et al. (2015) Challenges with the implementation of an Integrated Disease Surveillance and Response (IDSR) system: systematic review of the lessons learned. Health Policy Plan 30: 131-143. doi: 10.1093/heapol/czt097
|
| [67] |
Wölfel R, Stoecker K, Fleischmann E, et al. (2015) Mobile diagnostics in outbreak response, not only for ebola: A blueprint for a modular and robust field laboratory. Eurosurveillance 20: 30055. doi: 10.2807/1560-7917.ES.2015.20.44.30055
|
| [68] |
Kouadio K, Okeibunor J, Nsubuga P, et al. (2016) Polio infrastructure strengthened disease outbreak preparedness and response in the WHO African Region. Vaccine 34: 5175-5180. doi: 10.1016/j.vaccine.2016.05.070
|
| [69] |
Kruk ME (2008) Emergency Preparedness and Public Health Systems. Lessons for Developing Countries. Am J Prev Med 34: 529-534. doi: 10.1016/j.amepre.2008.02.012
|
| [70] |
Hulland EN, Wiens KE, Shirude S, et al. (2019) Travel time to health facilities in areas of outbreak potential: Maps for guiding local preparedness and response. BMC Med 17: 1-16. doi: 10.1186/s12916-019-1459-6
|
| [71] |
De Rooij D, Belfroid E, Eilers R, et al. (2020) Qualitative Research: Institutional Preparedness During Threats of Infectious Disease Outbreaks. BioMed Res Int 2020. doi: 10.1155/2020/5861894
|
| [72] |
Matua GA, Van der Wal DM, Locsin RC (2015) Ebola hemorrhagic fever outbreaks: Strategies for effective epidemic management, containment and control. Braz J Infect Dis 19: 308-313. doi: 10.1016/j.bjid.2015.02.004
|
| [73] |
Annan AA, Yar DD, Owusu M, et al. (2017) Health care workers indicate ill preparedness for Ebola Virus Disease outbreak in Ashanti Region of Ghana. BMC Public Health 17: 546. doi: 10.1186/s12889-017-4474-6
|
| [74] |
Omole O, Welye H, Abimbola S (2015) Boko Haram insurgency: Implications for public health. Lancet 385: 941. doi: 10.1016/S0140-6736(15)60207-0
|
| [75] |
Burki T (2016) Health crisis intensifying in Nigeria's Borno State. Vol. 388, Lancet (London, England). Lancet 388: 645. doi: 10.1016/S0140-6736(16)31345-9
|
| [76] |
Okunromade OF, Lokossou VK, Anya I, et al. (2019) Performance of the Public Health System during a Full-Scale Yellow Fever Simulation Exercise in Lagos State, Nigeria, in 2018: How Prepared Are We for the Next Outbreak. Heal Secur 17: 485-494. doi: 10.1089/hs.2019.0048
|
| [77] | NCDC (2018) President Muhammadu Buhari Signs Bill For An Act To Establish The NCDC. Available from: https://www.ncdc.gov.ng/news/156/president-muhammadu-buhari-signs-bill-for-an-act-to-establish-the-ncdc. |
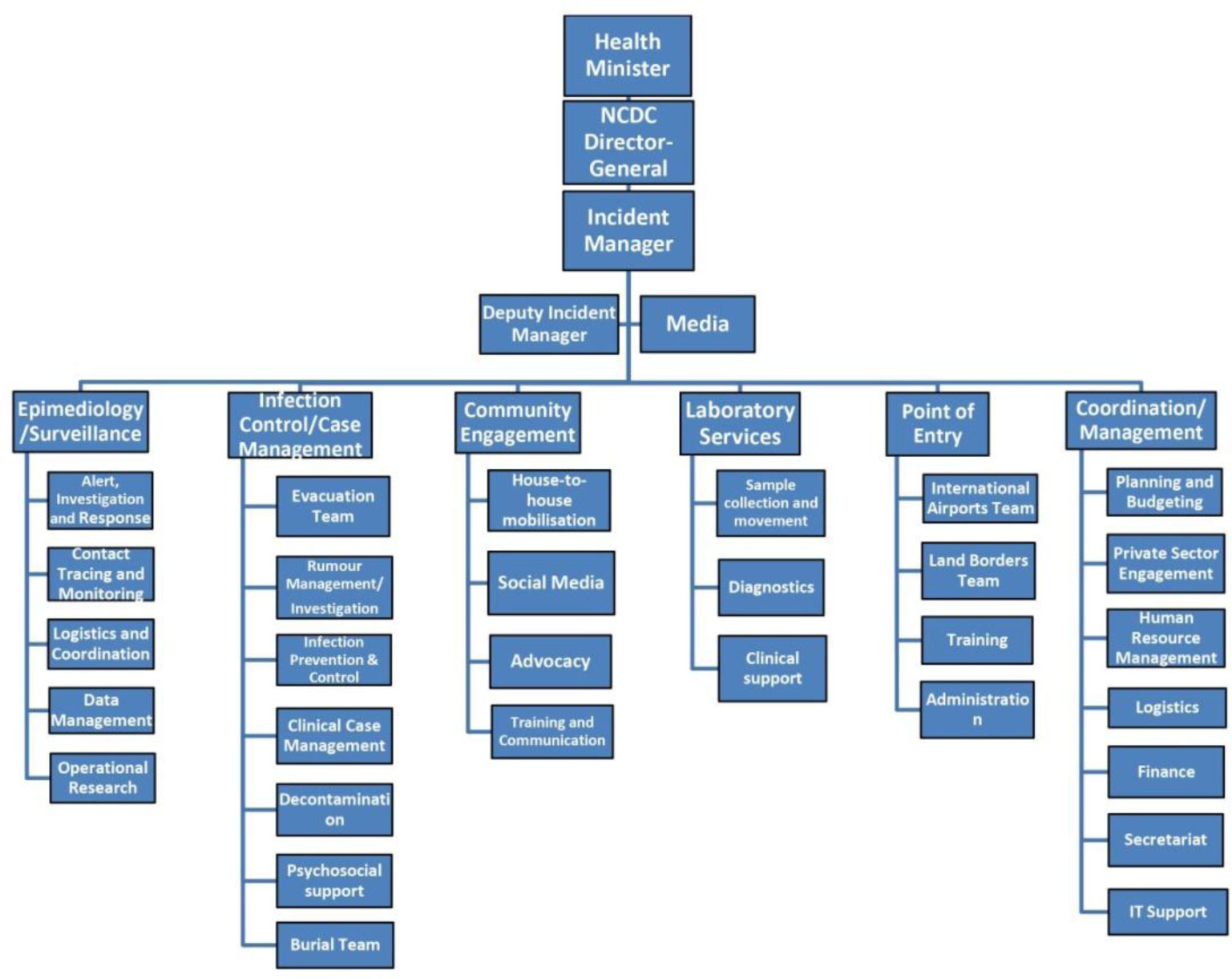
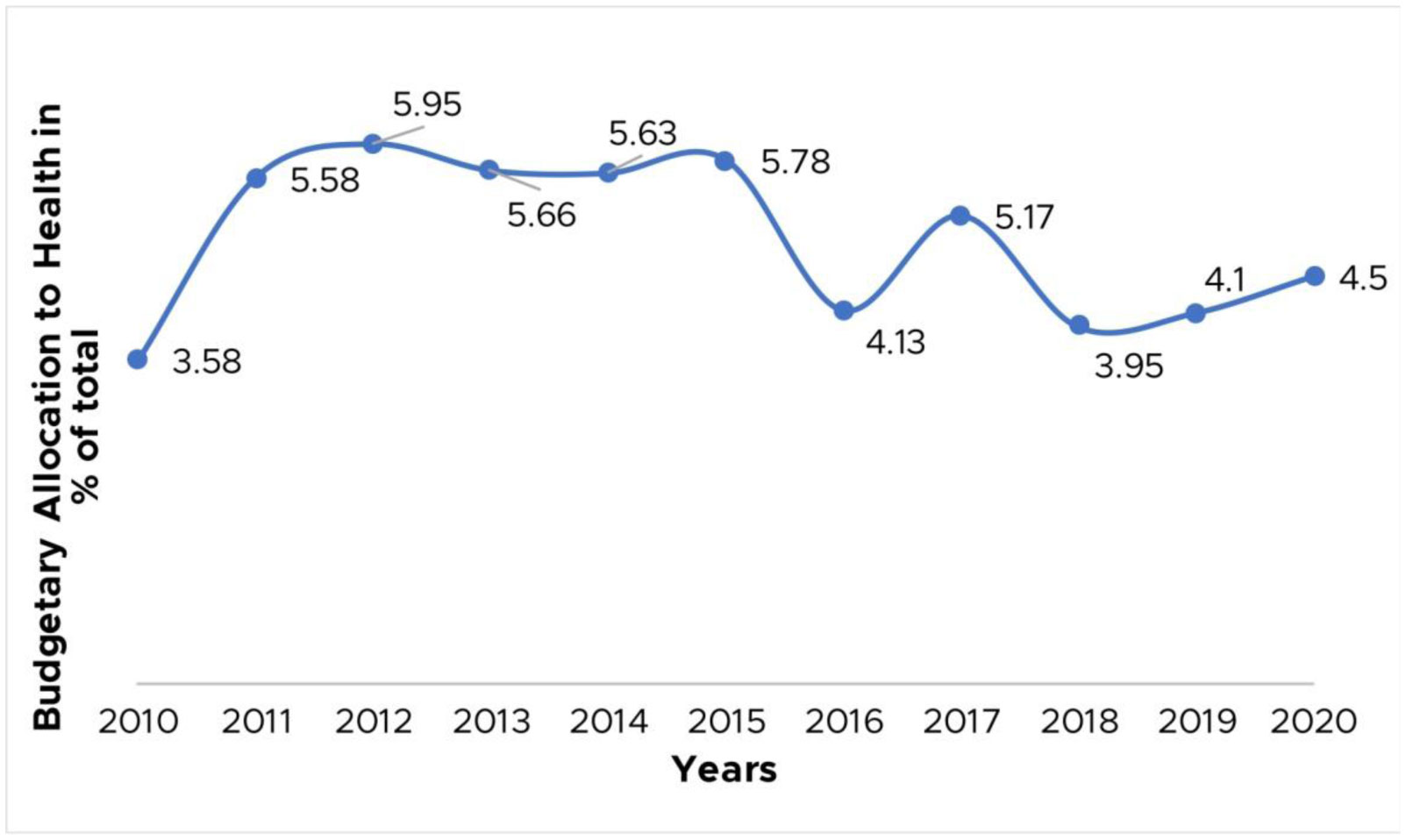
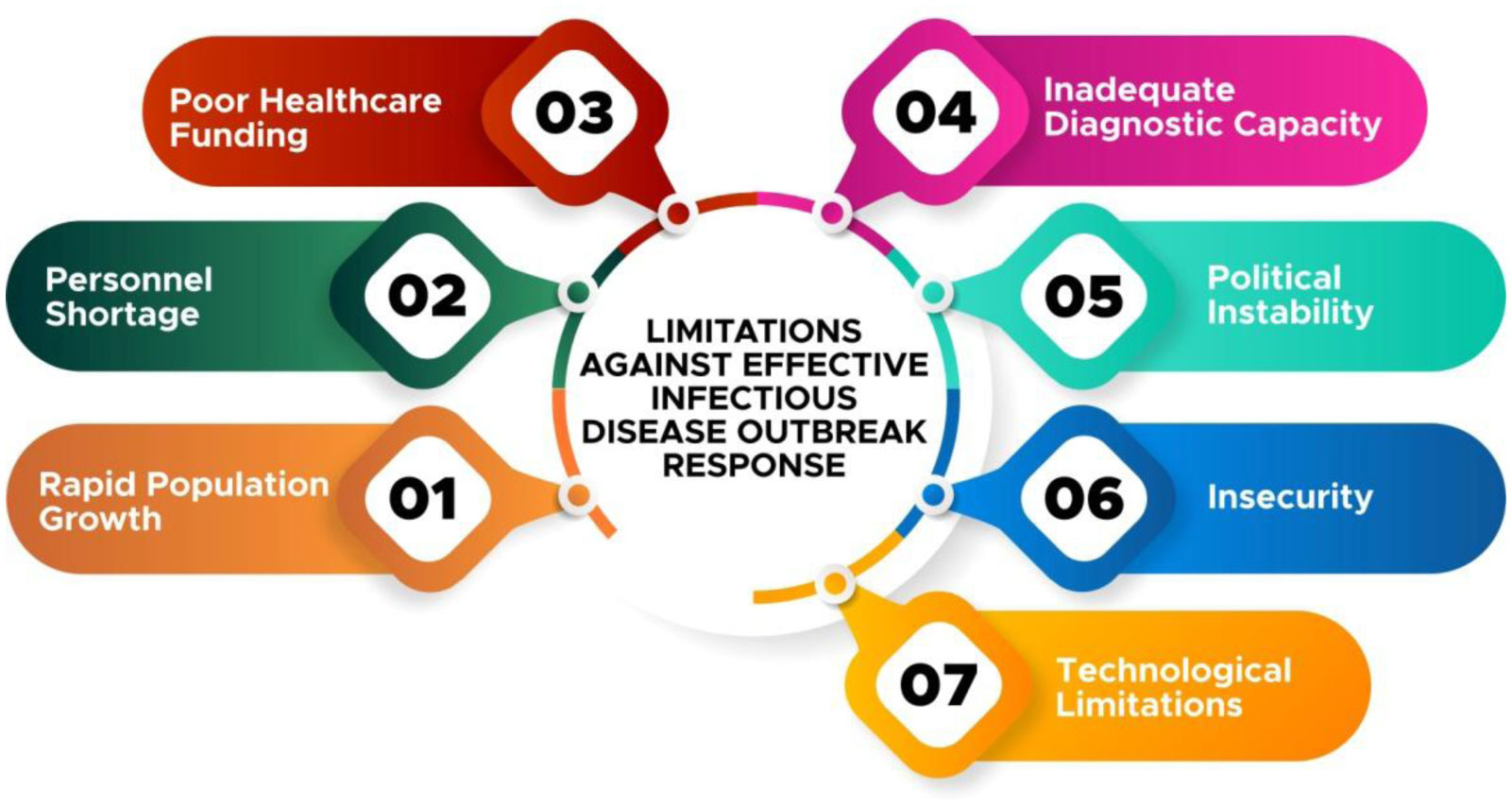
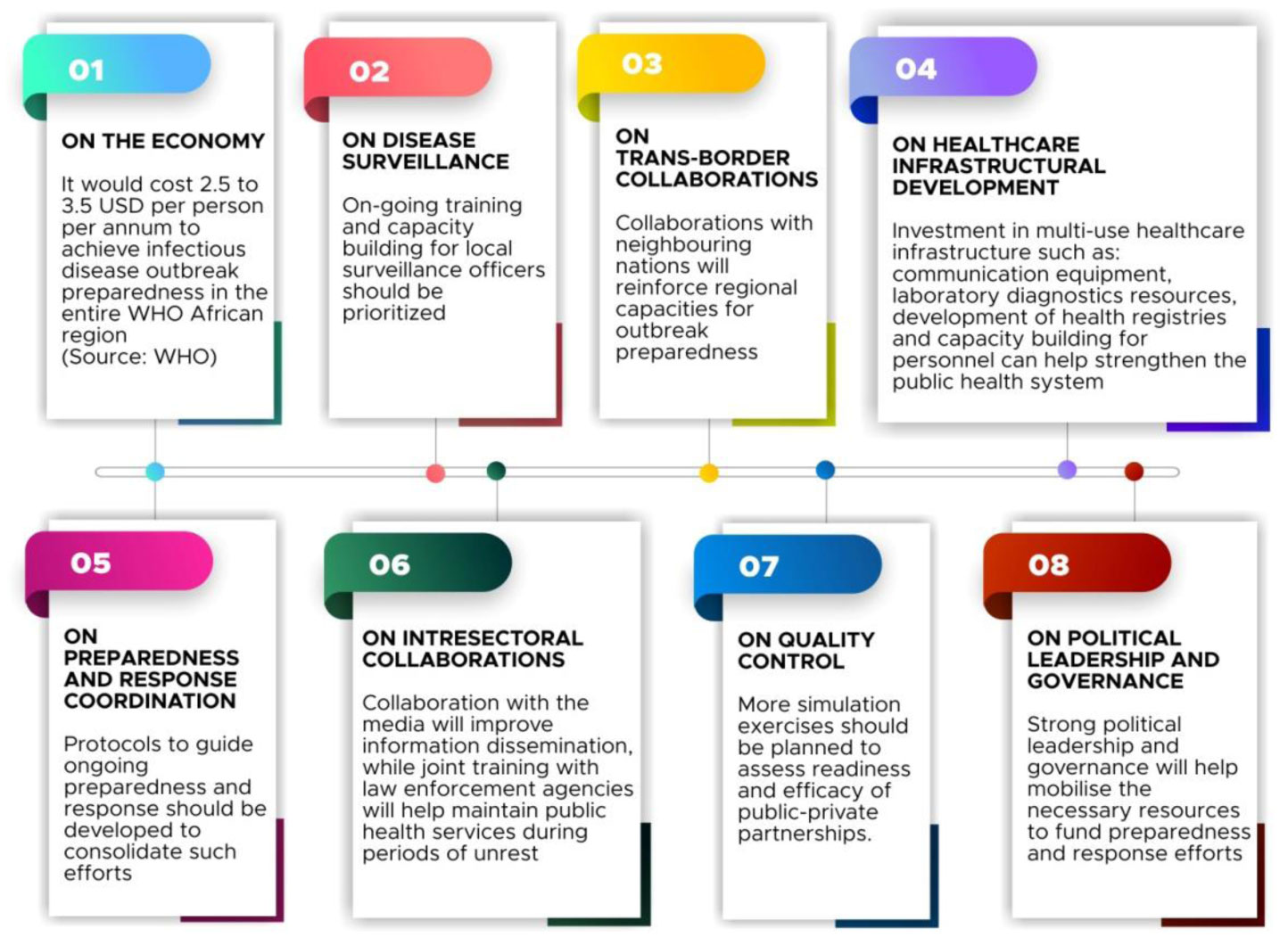
Figures(4)

Testimony J Olumade, Oluwafolajimi A Adesanya, Iyanuoluwa J Fred-Akintunwa, David O Babalola, Judith U Oguzie, Olusola A Ogunsanya, Uwem E George, Oluwawapelumi D Akin-Ajani, Damilola G Osasona. Infectious disease outbreak preparedness and response in Nigeria: history, limitations and recommendations for global health policy and practice[J]. AIMS Public Health, 2020, 7(4): 736-757. doi: 10.3934/publichealth.2020057







 DownLoad:
DownLoad: