Citation: Marcos André dos Santos, Alfredo Jarma-Orozco, Flavio Lozano-Isla, José Nailson S. Barros, Jesús Rivera, Carlos Espitia-Romero, Ángela Castillejo-Morales, Betty Jarma-Arroyo, Marcelo Francisco Pompelli. Leaf area estimation in Jatropha curcas (L.): an update[J]. AIMS Environmental Science, 2018, 5(5): 353-371. doi: 10.3934/environsci.2018.5.353

| [1] | Pecina-Quintero V, Anaya-López JL, Zamarripa-Colmenero A, et al. (2014) Genetic structure of Jatropha curcas L. in Mexico and probable centre of origin. Biomass Bioenerg 60: 147–155. |
| [2] |
Silitonga AS, Hassan MH, Ong HC, et al. (2017) Analysis of the performance, emission and combustion characteristics of a turbocharged diesel engine fuelled with Jatropha curcas biodiesel-diesel blends using kernel-based extreme learning machine. Environ Sci Pollut Res 24: 25383–25405. doi: 10.1007/s11356-017-0141-9

|
| [3] |
Mardhiah HH, Ong HC, Masjuki HH, et al. (2017) Investigation of carbon-based solid acid catalyst from Jatropha curcas biomass in biodiesel production. Energy Convers Manag 144: 10–17. doi: 10.1016/j.enconman.2017.04.038
|
| [4] |
Alburquerque N, García-Almodóvar RC, Valverde JM, et al. (2017) Characterization of Jatropha curcas accessions based in plant growth traits and oil quality. Ind Crops Prod 109: 693–698. doi: 10.1016/j.indcrop.2017.09.019
|
| [5] | Corte-Real N, Endres L, Santos KPO, et al. (2016) Morphoanatomy and ontogeny of the fruit and seeds of Jatropha curcas L.: A promising biofuel plant. In: Segura-Campos MR, Betancur-Ancova D, editors. The Promising Future of Jatropha curcas: Proprieties and potential applications. Hauppauge, NY: Nova Science Publishers, Inc. pp. 141–158. |
| [6] | Heller J (1996) Physic nut. Jatropha curcas L. Promoting the conservation and use of underutilized and neglected crops. Rome: International Plant Genetic Resources Institute (IPGRI). 66 p. |
| [7] |
Pandey VC, Singh K, Singh JS, et al. (2012) Jatropha curcas: A potential biofuel plant for sustainable environmental development. Renew Sust Energy Rev 16: 2870–2883. doi: 10.1016/j.rser.2012.02.004
|
| [8] |
Pompelli MF, Ferreira DTRG, Cavalcante PPGS, et al. (2010) Environmental influence on the physico-chemical and physiological properties of Jatropha curcas L. seeds. Aust J Bot 58: 421–427. doi: 10.1071/BT10102
|
| [9] |
Contran N, Chessa L, Lubino M, et al. (2013) State-of-the art of the Jatropha curcas productive chain: From sowing to biodiesel and by-products. Ind Crops Prod 42: 202–215. doi: 10.1016/j.indcrop.2012.05.037
|
| [10] |
Divakara BN, Upadhyaya HD, Wani SP, et al. (2010) Biology and genetic improvement of Jatropha curcas L. A review. Appl Energ 87: 732–742. doi: 10.1016/j.apenergy.2009.07.013
|
| [11] | Pompelli MF, Orozco AJ, Oliveira MTO, et al. (2011) Crise energética mundial e o papel do Brasil na problemática de biocombustíveis. Agronomía Colombiana 29: 361–371. |
| [12] |
Openshaw K (2000) A review of Jatropha curcas: an oil plant of unfulfilled promise. Biomass Bioenerg 19: 1–15. doi: 10.1016/S0961-9534(00)00019-2
|
| [13] |
Chel A, Kaushik G (2011) Renewable energy for sustainable agriculture. Agron Sustain Dev 31: 91–118. doi: 10.1051/agro/2010029
|
| [14] |
Gutiérrez-Antonio C, Romero-Izquierdo AG, Gómez-Castro FI, et al. (2016) Simultaneous energy integration and intensification of the hydrotreating process to produce biojet fuel from Jatropha curcas. Chem Eng Process 110: 134–145. doi: 10.1016/j.cep.2016.10.007
|
| [15] | Beaver A, Castaño AG, Díaz MS (2016) Life cycle analysis of Jatropha curcas as a sustainable biodiesel feedstock in Argentina. Chem Engin Trans 50. |
| [16] |
Galati A, Crescimanno M, Gristina L, et al. (2016) Actual provision as an alternative criterion to improve the efficiency of payments for ecosystem services for C sequestration in semiarid vineyards. Agric Systems 144: 58–64. doi: 10.1016/j.agsy.2016.02.004
|
| [17] |
Keesstra SD, Bouma J, Wallinga J, et al. (2016) The significance of soils and soil science towards realization of the United Nations Sustainable Development Goals. Soil 2: 111–128. doi: 10.5194/soil-2-111-2016
|
| [18] | Brittaine R, Lutaladio N (2010) Jatropha: A Smallholder Bioenergy Crop The Potential for Pro-Poor Development. Rome Italy: Plant Production and Protection Division FAO. 96 p. |
| [19] |
Díaz S, Kattge J, Cornelissen JHC, et al. (2016) The global spectrum of plant form and function. Nature 529: 167–171. doi: 10.1038/nature16489
|
| [20] |
Cristofori V, Rouphael Y, Mendoza-de Gyves E, et al. (2007) A simple model for estimating leaf area of hazelnut from linear measurements. Sci Hort 113: 221–225. doi: 10.1016/j.scienta.2007.02.006
|
| [21] |
Pompelli MF, Antunes WC, Ferreira DTRG, et al. (2012) Allometric models for non-destructive leaf area estimation of the Jatropha curcas. Biomass Bioenerg 36: 77–85. doi: 10.1016/j.biombioe.2011.10.010
|
| [22] |
Blanco FF, Folegatti MV (2005) Estimation of leaf area for greenhouse cucumber by linear measurements under salinity and grafting. Sci Agr 62: 305–309. doi: 10.1590/S0103-90162005000400001
|
| [23] |
Antunes WC, Pompelli MF, Carretero DM, et al. (2008) Allometric models for non-destructive leaf area estimation in coffee (Coffea arabica and Coffea canephora). Ann Appl Biol 153: 33–40. doi: 10.1111/j.1744-7348.2008.00235.x
|
| [24] |
Keramatlou I, Sharifani M, Sabouri H, et al. (2015) A simple linear model for leaf area estimation in Persian walnut (Juglans regia L.). Sci Hort 184: 36–39. doi: 10.1016/j.scienta.2014.12.017
|
| [25] |
Liu Z, Zhu Y, Li F, et al. (2017) Non-destructively predicting leaf area, leaf mass and specific leaf area based on a linear mixed-effect model for broadleaf species. Ecol Indic 78: 340–350. doi: 10.1016/j.ecolind.2017.03.025
|
| [26] |
Peksen E (2007) Non-destructive leaf area estimation model for faba bean (Vicia faba L.). Sci Hortic-Amsterdam 113: 322–328. doi: 10.1016/j.scienta.2007.04.003
|
| [27] | Thomas B (2017) Leaf Development. In: Thomas B, Murray BG, Murphy DJ, editors. Encyclopedia of Applied Plant Sciences (Second Edition). San Diego: Elsevier. pp. 191–197. |
| [28] |
Kirkman LK, Sharitz RR (1994) Vegetation disturbance and maintenance of diversity in intermittently flooded Carolina bays in South Carolina. Ecol Appl 4: 177–188. doi: 10.2307/1942127
|
| [29] |
Collins SL (1987) Interaction of disturbance in tallgrass prairie: a field experiment. Ecology 68: 1243–1250. doi: 10.2307/1939208
|
| [30] |
Steel M, Penny D (2000) Parsimony, likelihood, and the role of models in molecular phylogenetics. Mol Biol Evol 17: 839–850. doi: 10.1093/oxfordjournals.molbev.a026364
|
| [31] |
Cumming G, Fidler F, Vaux DL (2007) Error bars in experimental biology. J Cell Biol 177: 7–11. doi: 10.1083/jcb.200611141
|
| [32] |
Walther BA, Moore JL (2005) The concepts of bias, precision and accuracy, and their use in testing the performance of species richness estimators, with a literature review of estimator performance. Ecography 28: 815–829. doi: 10.1111/j.2005.0906-7590.04112.x
|
| [33] | Messier J, McGill BJ, Lechowicz MJ (2010) Ecol Lett 13: 838–848. |
| [34] | Pinheiro J, Bates D, DebRoy S, et al. (2017) nlme:Linear and Nonlinear Mixed Effects Models. R package version 3.1–131. Viena, Austria: R Foundation for Statistical Computing. |
| [35] | Achten WMJ, Maes WH, Reubens B, et al. (2010) Biomass production and allocation in Jatropha curcas L. seedlings under different levels of drought stress. Biomass Bioenerg 34: 667–676. |
| [36] | Severino LS, Vale LS, Beltrão NEM (2007) A simple method for measurement of Jatropha curcas leaf area. Rev Bras Ol Fibros 11: 9–14. |
| [37] | Ahmed N, Khan D (2011) Leaf area estimation in Jatropha curcas L. Int J Biol Biotech 8: 401–407. |
| [38] |
Zuur AF, Elena NI, Elphick CS (2010) A protocol for data exploration to avoid common statistical problems. Method Ecol Evol 1: 3–14. doi: 10.1111/j.2041-210X.2009.00001.x
|
| [39] |
Kandiannan K, Parthasarathy U, Krishnamurthy KS, et al. (2009) Modeling individual leaf area of ginger (Zingiber officinale Roscoe) using leaf length and width. Sci Hortic-Amsterdam 120: 532–537. doi: 10.1016/j.scienta.2008.11.037
|
| [40] |
Souza MC, Amaral CL (2015) Non-destructive linear model for leaf area estimation in Vernonia ferruginea Less. Braz J Biol 75: 152–156. doi: 10.1590/1519-6984.09813
|
| [41] |
Villar R, Ruiz-Robleto J, Ubera JL, et al. (2013) Exploring variation in leaf mass per area (LMA) from leaf to cell: an anatomical analysis of 26 woody species. Am J Bot 100: 1969–1980. doi: 10.3732/ajb.1200562
|
| [42] |
Zhang L, Pan L (2011) Allometric models for leaf area estimation across different leaf-age groups of evergreen broadleaved trees in a subtropical forest. Photosynthetica 49: 219–226. doi: 10.1007/s11099-011-0027-x
|
| [43] |
Tondjo K, Brancheriau L, Sabatier SA, et al. (2015) Non-destructive measurement of leaf area and dry biomass in Tectona grandis. Trees 29: 1625–1631. doi: 10.1007/s00468-015-1227-y
|
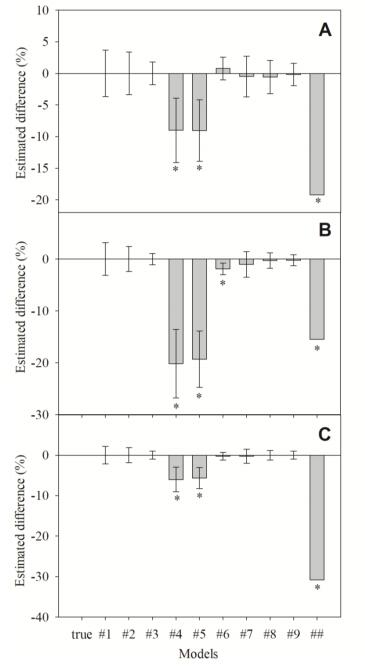
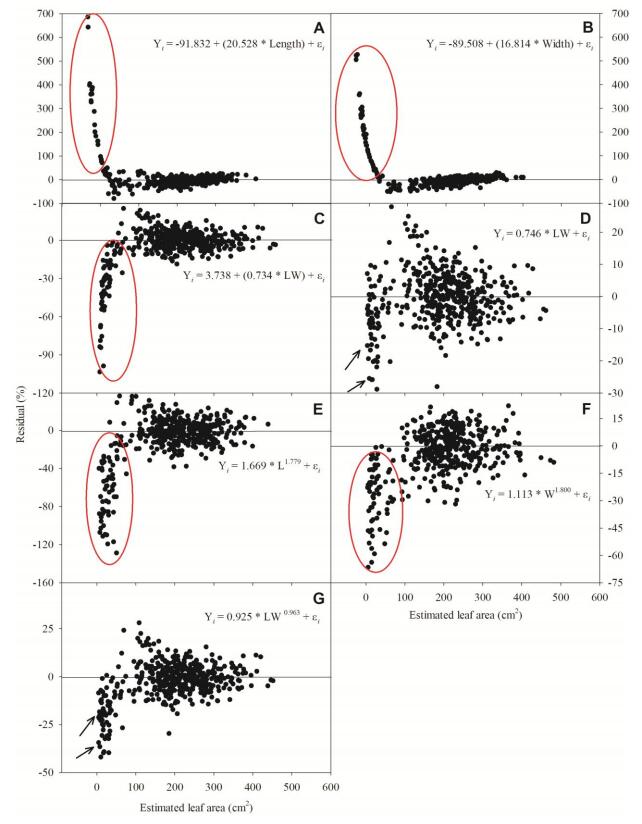
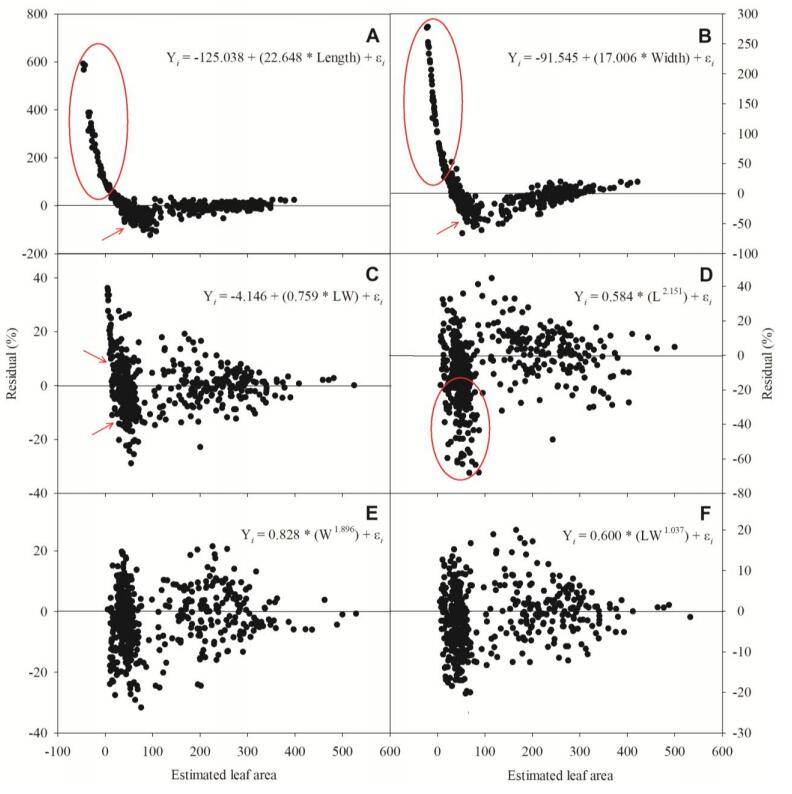
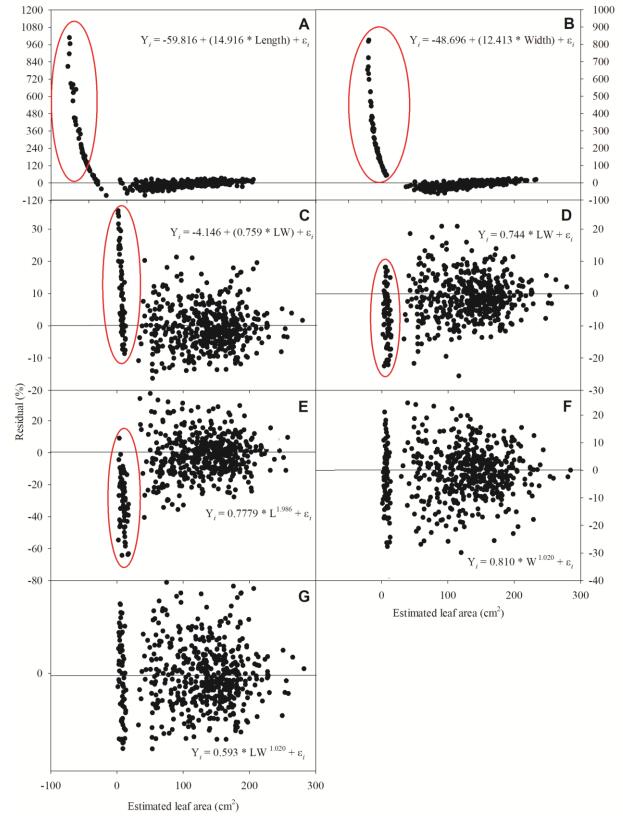
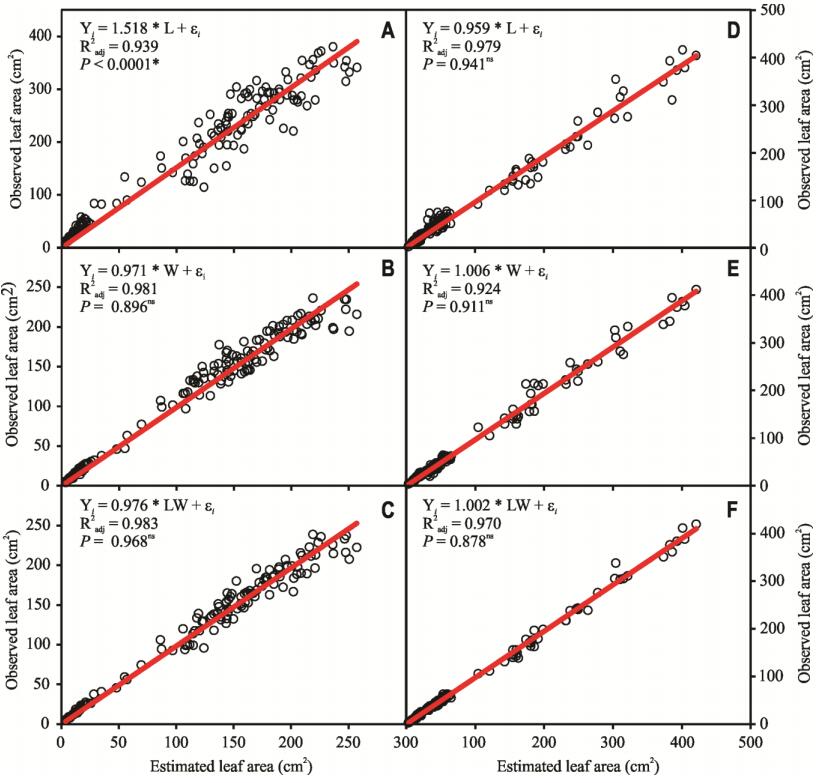
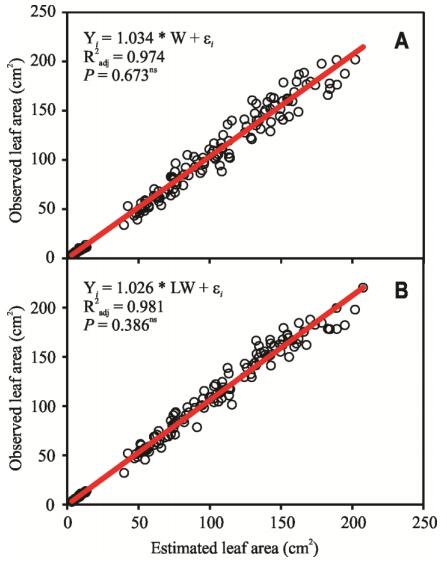
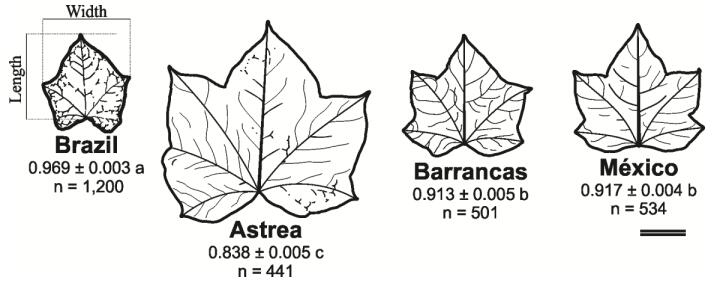
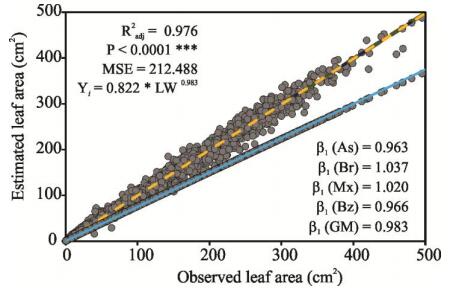
Figures(8) / Tables(4)

Marcos André dos Santos, Alfredo Jarma-Orozco, Flavio Lozano-Isla, José Nailson S. Barros, Jesús Rivera, Carlos Espitia-Romero, Ángela Castillejo-Morales, Betty Jarma-Arroyo, Marcelo Francisco Pompelli. Leaf area estimation in Jatropha curcas (L.): an update[J]. AIMS Environmental Science, 2018, 5(5): 353-371. doi: 10.3934/environsci.2018.5.353







 DownLoad:
DownLoad: