Citation: Elizabeth Ramírez-Iglesias, Rosa M. Hernández-Hernández, Carlos Bravo, José R. Ramírez-Iglesias, Pablo Herrera. Consumption, digestibility and cattle condition according to forage composition and quality in maize-cattle from conservation-based agroecosystems[J]. AIMS Agriculture and Food, 2020, 5(3): 480-499. doi: 10.3934/agrfood.2020.3.480

| [1] | Herrera P, Birbe B, Colmenares O, et al. (2008) Sistemas de producción con ganadería de doble propósito en condiciones de sabanas bien drenadas. Acta Biol Venez 28: 29-38. |
| [2] | Mendoza JC, Aparicio YV, Jerez MP, et al. (2013) Ecological evaluation of three agroecosystems of sheep production in Central Valleys of Oaxaca. Rev Mex Cienc Agr 6: 1251-1261. |
| [3] | Clements RJ (1996) Pastures for prosperity. 3. The future for new tropical pasture plants. Trop Grasslands 30: 31-46. |
| [4] |
Nakamura T, Miranda CH, Ohwaki Y, et al. (2005) Characterization of nitrogen utilization by Brachiaria grasses in Brazilian Savannas (Cerrados). Soil Sci Plant Nutr 51: 973-979. doi: 10.1111/j.1747-0765.2005.tb00136.x

|
| [5] | Vercoe JE (1996) Pastures for prosperity. 5. The role of animal science. Trop Grasslands 30: 58-72. |
| [6] | Hernández-Hernández RM, Lozano Z, Toro M, et al. (2011) Informe final de proyecto: "Manejo agroecológico de suelos de sabanas bien drenadas con unidades de producción cereal-ganado." FONACIT-MPPCyT. Caracas, Venezuela, 202. |
| [7] |
Benvenutti MA, Pavetti DR, Poppi DP, et al. (2016) Defoliation patterns and their implications for the management of vegetative tropical pastures to control intake and diet quality by cattle. Grass Forage Sci 71: 424-436. doi: 10.1111/gfs.12186
|
| [8] | Barriga J, Visbal M, Acero J (2011) Relación entre los caracteres de las micorrizas arbusculares nativas con las propiedades físico-químicas del suelo y bromatología del pasto estrella en ganadería de carne. Rev Científica UDO Agr 11: 134-141. |
| [9] |
Pedreira CG, Braga GJ, Portela JN (2017) Herbage accumulation, plant-part composition and nutritive value on grazed signal grass (Brachiaria decumbens) pastures in response to stubble height and rest period based on canopy light interception. Crop Pasture Sci 68: 62-73. doi: 10.1071/CP16333
|
| [10] |
Trevisi E, Riva F, Filipe JFS, et al. (2018) Innate immune responses to metabolic stress can be detected in rumen fluids. Res Vet Sci 117: 65-73. doi: 10.1016/j.rvsc.2017.11.008
|
| [11] |
Cuchillo-Hilario M, Wrage-Mönnig N, Isselstein J (2018) Forage selectivity by cattle and sheep co-grazing swards differing in plant species diversity. Grass Forage Sci 73: 320-329. doi: 10.1111/gfs.12339
|
| [12] |
Couto RK, Pinto GG, Rodrigues R, et al. (2016) Comparison of protein and energy supplementation to mineral supplementation on feeding behavior of grazing cattle during the rainy to the dry season transition. Springerplus 5: 2-7. doi: 10.1186/s40064-015-1651-x
|
| [13] |
Maestri R, Monteiro LR, Fornel R, et al. (2018) Geometric morphometrics meets metacommunity ecology: environment and lineage distribution affects spatial variation in shape. Ecography 41: 90-100. doi: 10.1111/ecog.03001
|
| [14] | Anza M, Epelde L, Mijangos I, et al. (2016) Effects of grazing abandonment on the health of pasture ecosystems. Pastos 45: 15-22. |
| [15] |
Vélez OM, Campos R, Sánchez H, et al. (2017) Evaluación de diferentes niveles de inclusión de plantas nativas de sabanas inundables sobre una dieta basal de Brachiaria humidicola y su efecto sobre la producción de metano in vitro. Arch Zootec 66: 341-349. doi: 10.21071/az.v66i255.2509
|
| [16] |
DePeters EJ, George LW (2014) Rumen transfaunation. Immunol Lett 162: 69-76. doi: 10.1016/j.imlet.2014.05.009
|
| [17] | Smith LW, Goering HK, Gordon CH (1969) Influence of chemical treatments upon digestibility of ruminant feces. Proc Conf on Anim Waste Manage 88-97. |
| [18] | Caballero JC, Moreno A, Reyes JL, et al. (2017) Competencia del uso del rastrojo de maíz en sistemas agropecuarios mixtos en Chiapas. Rev Mex Cienc Agr 8: 91-104. |
| [19] |
Orgill SE, Condon JR, Conyers MK, et al. (2018) Removing grazing pressure from a native pasture decreases soil organic carbon in southern New South Wales, Australia. Land Degrad Dev 29: 274-283. doi: 10.1002/ldr.2560
|
| [20] | Ormaechea S, Gargaglione V, Bahamonde HA, et al. (2018) Producción bovina bajo manejo silvopastoril intensivo a escala de establecimiento y ciclo completo en Tierra del Fuego, Argentina. Livest Res Rural Dev 30: 1-12. |
| [21] | Gonçalves Z, Junqueira EC, Pinto F, et al. (2019) Morphogenic, structural, productive and bromatological characteristics of Braquiária in silvopastoral system under nitrogen doses. Acta Sci Anim Sci 41: 1-8. |
| [22] | Santos BRC dos, Alfaya H, Dias AEA, et al. (2011) Correlation of the phosphorus in the system soil-plant-animal in natural pasture in the region of campanha-rs. Arch Zootec 60: 487-497. |
| [23] | Givens DI, Owen E, Omed HM, et al. (2000) Forage Evaluation in Ruminant Nutrition. CABI, Wallingford, UK, 498. |
| [24] | Ginantra IK, Made IB, Muksin IK (2018) Selection of forages by timor deer (Cervus timorensis blainville) in Menjangan island, Bali. IOP Conf Ser Earth Environ Sci 130: 1-12. |
| [25] | Sparks DR, Malechek JC (1968) Estimating percentage dry weight in diets using a microscopic technique. Rangeland Ecol Manag/J Range Manag Arch 21: 264-265. |
| [26] |
Tóth E, Deák B, Valkó O, et al. (2018) Livestock type is more crucial than grazing intensity: traditional cattle and sheep grazing in short-grass steppes. Land Degrad Dev 29: 231-239. doi: 10.1002/ldr.2514
|
| [27] | Mata D, Herrera P, Birbe B (1996) Sistemas de producción animal con bajos insumos para las sabanas de Trachypogon sp. Ecotropicos 9: 83-100. |
| [28] | Ramia M (1967) Tipos de sabanas en los llanos de Venezuela. Bol Soc Ven Cienc Nat 27: 264-288. |
| [29] | Hernández-Hernández RM, Florentino A, López-Hernández D (2000) Efectos de la siembra directa y la labranza convencional sobre la estabilidad estructural y otras propiedades físicas de un suelo de sabana. Agron Trop 50: 9-29. |
| [30] | Carpenter SR (1998) The Need for Large-Scale Experiments to Assess and Predict the Response of Ecosystems to Perturbation, In: Pace ML, Groffman PM, editors. Successes, Limitations, and Frontiers in Ecosystem Science, New York, NY: Springer, 287-312. |
| [31] | Lozano Z, Bravo C, Ovalles F, et al. (2004) Selección de un diseño de muestreo en parcelas experimentales a partir del estudio de la variabilidad espacial de los suelos. Bioagro 16: 61-72. |
| [32] | AOAC (1998) Methods of analysis (16th). Washington D.C. EUA. |
| [33] | Brown S (1997) Estimating Biomass and Biomass Change of Tropical Forests: A Primer. FAO Forestry Paper, Roma, Italy, 134. |
| [34] | Sepúlveda L, Pelliza A, Manacorda M (2004) La importancia de los tejidos no epidérmicos en el microanálisis de la dieta de herbívoros. Ecol Austral 14: 31-38. |
| [35] | Castellaro G, Ullrich T, Wackwitz B, et al. (2004) Composición botánica de la dieta de alpacas (Lama pacos L.) y llamas (Lama glama L.) en dos estaciones del año, en praderas altiplánicas de un sector de la Provincia de Parinacota, Chile. Agr Técnica 64: 353-363. |
| [36] |
Holechek JL, Vavra M, Pieper RD (1982) Botanical composition determination of range herbivore diets: a review. J Range Manag 35: 309-315. doi: 10.2307/3898308
|
| [37] |
Morante N, Sánchez T, Ceballos H, et al. (2010) Tolerance to postharvest physiological deterioration in cassava roots. Crop Sci 50: 1333-1338. doi: 10.2135/cropsci2009.11.0666
|
| [38] | Soest PJV (2018) Nutritional Ecology of the Ruminant. Ithaca, NY: Cornell University Press, 490. |
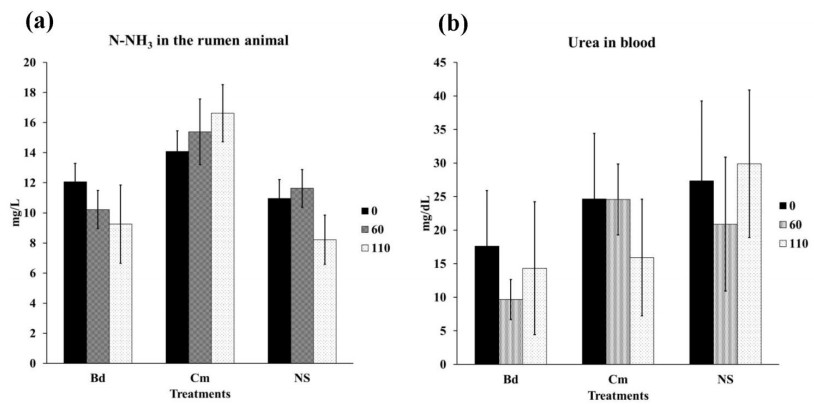
| [39] | Pardo O, Carulla JE, Hess HD (2008) Efecto de la relación proteína y energía sobre los niveles de amonio ruminal y nitrógeno ureico en sangre y leche de vacas doble propósito del piedemonte llanero, Colombia. Rev Col Cienc Pecuarias 21: 387-397. |
| [40] | Di Rienzo JA, Casanoves F, Balzarini MG, et al. (2011) Grupo InfoStat FCA, Universidad Nacional de Córdoba, Argentina. 8: 195-199. |
| [41] | Uriel E, Manzano JA (2002) Análisis multivariante aplicado. Paraninfo 76: 270-271. |
| [42] | Ramírez-Iglesias E, Hernández-Hernández RM, Herrera P (2017) Relaciones suelo-planta-animal en un sistema agroecológico de siembra directa y asociación de coberturas maíz-ganado en sabanas bien drenadas de Venezuela. Acta Biol Venez 37: 67-87. |
| [43] | Rodas-González A, Vergara-López J, Arenas L, et al. (2006) Características al sacrificio, rasgos de la canal y rendimiento carnicero de novillos criollo limonero sometidos a suplementación durante la fase de ceba a pastoreo. Rev Científica 16: 315-324. |
| [44] |
Carvalho ÁC, Pereira M, Bonomo P, et al. (2014) Microbial protein synthesis and nitrogen metabolism in cows bred on tropical pasture and fed on cassava root and corn. Acta Sci Anim Sci 36: 185-192. doi: 10.4025/actascianimsci.v36i2.22161
|
| [45] | Russell JB, O'Connor JD, Fox DG, et al. (1992) A net carbohydrate and protein system for evaluating cattle diets: I. Ruminal fermentation. J Anim Sci 70: 3551-3561. |
| [46] |
Pfeiffer M, Langan L, Linstädter A, et al. (2019) Grazing and aridity reduce perennial grass abundance in semi-arid rangelands - Insights from a trait-based dynamic vegetation model. Ecol Model 395: 11-22. doi: 10.1016/j.ecolmodel.2018.12.013
|
| [47] | Sevim Ö, Önol AG (2019) Supplemental slow-release urea and non-structural carbohydrates: effect on digestibility and some rumen parameters of sheep and goats. J Anim Plant Sci 29: 1-7. |
| [48] | Ramírez-Iglesias E, Hernández-Hernández RM, Herrera P (2017) Dinámica del fósforo en un agroecosistema maíz-ganado de sabanas neotropicales. Rev Col Cienc Anim Recia 9: 147-157. |
| [49] | La O, González H, Orozco A, et al. (2012) Composición química, degradabilidad ruminal in situ y digestibilidad in vitro de ecotipos de Tithonia diversifolia de interés para la alimentación de rumiantes. Rev Cubana Cienc Agr 46: 47-53. |
| [50] | Singh S, Nag SK, Kundu SS, et al. (2010) Relative intake, eating pattern, nutrient digestibility, nitrogen metabolism, fermentation pattern and growth performance of lambs fed organically and inorganically produced cowpea hay- barley grain diets. Trop Grasslands 44: 55-61. |
| [51] | Giráldez FJ, Mantecón ÁR, Chaso MA, et al. (1994) Efecto del tipo de dieta y de la frecuencia de alimentación sobre la actividad degradativa ruminal. Prod Sanid Anim 9: 245-259. |
| [52] |
Satter LD, Slyter LL (1974) Effect of ammonia concentration on rumen microbial protein production in vitro. British J Nutr 32: 199-208. doi: 10.1079/BJN19740073
|
| [53] | Chamorro DR, Carulla JE, Cuesta PA (2005) Caracterización nutricional de dos asociaciones gramínea-leguminosa con novillas en pastoreo en el Alto Magdalena. Ciencia y Tecnología Agropecuaria 6: 37-51. |
| [54] | Sales E, Varela AM, Ferreira L, et al. (2005) Importância da inter-relação carboidrato e proteína em dietas de ruminantes. Sem Ci Agr 30: 125-134. |
| [55] | Bravo C, Lozano Z, Hernández-Hernández RM, et al. (2004) Efecto de diferentes especies de coberturas sobre las propiedades físicas de un suelo de sabana con siembra directa de maíz. Bioagro 16: 163-172. |
| [56] | Lozano Z, Hernández-Hernández RM, Bravo C, et al. (2012) Disponibilidad de fósforo en un suelo de las sabanas bien drenadas venezolanas, bajo diferentes tipos de cobertura y fuentes de fertilización. Interciencias 37: 820-827. |
| [57] | Canelones C, Castejon M (2006) Flours of whole cowpea (Vigna unguiculata) and cob maize (Zea mays) as supplements for suckling calves. Zootec Trop 24: 361-378. |
| [58] | Milligan KE, Brookes IM, Thompson KF (1987) Feed planning on pasture, In: Nicol, A.M., Livestock feeding on pasture, New Zealand Society of Animal Production: Hamilton, 75-88. |
| [59] | Thiago LRL, Gill M (1990) Consumo voluntário: fatores relacionados com a degradação e passagem da forragem pelo rumen. EMBRAPA, Campo Grande, Brasil, 65. |

Figures(5) / Tables(3)

Elizabeth Ramírez-Iglesias, Rosa M. Hernández-Hernández, Carlos Bravo, José R. Ramírez-Iglesias, Pablo Herrera. Consumption, digestibility and cattle condition according to forage composition and quality in maize-cattle from conservation-based agroecosystems[J]. AIMS Agriculture and Food, 2020, 5(3): 480-499. doi: 10.3934/agrfood.2020.3.480







 DownLoad:
DownLoad: