Citation: Mohamed Amine Ben Mlouka, Thomas Cousseau, Patrick Di Martino. Application of fluorescently labelled lectins for the study of polysaccharides in biofilms with a focus on biofouling of nanofiltration membranes[J]. AIMS Molecular Science, 2016, 3(3): 338-356. doi: 10.3934/molsci.2016.3.338

| [1] | Mounier J, Camus A, Mitteau I, et al. (2014) The marine bacterium Marinobacter hydrocarbonoclasticus SP17 degrades a wide range of lipids and hydrocarbons through the formation of oleolytic biofilms with distinct gene expression profiles. FEMS Microbiol Ecol 90: 816–831. |
| [2] | Nait Chabane Y, Marti S, Rihouey C, et al. (2014) Characterisation of pellicles formed by Acinetobacter baumannii at the air-liquid interface. PloS One 9: e111660. |
| [3] |
Gagnière H, Di Martino P (2004) Effects of antibiotics on Pseudomonas aeruginosa NK125502 and Pseudomonas fluorescens MF0 biofilm formation on immobilized fibronectin. J Chemother Florence Italy 16: 244–247. doi: 10.1179/joc.2004.16.3.244

|
| [4] | Flemming HC, Schaule G (1996) Measures against biofouling. In: Heitz E, Sand W, Flemming H-C, Eds. Microbially influenced corrosion of materials – scientific and technological aspects. Springer, Berlin. 121–139. |
| [5] |
Di Martino P (2016) What About Biofilms on the Surface of Stone Monuments? Open Conf Proc J 6: 14–28. doi: 10.2174/2210289201607020014
|
| [6] |
Costerton JW, Stewart PS, Greenberg EP (1999) Bacterial biofilms: a common cause of persistent infections. Science 284: 1318–1322. doi: 10.1126/science.284.5418.1318
|
| [7] | Vastano V, Pagano A, Fusco A, et al. (2016) The Lactobacillus plantarum Eno A1 Enolase Is Involved in Immunostimulation of Caco-2 Cells and in Biofilm Development. Adv Exp Med Biol 897: 33–44. |
| [8] |
Klein B, Grossi V, Bouriat P, et al. (2008) Cytoplasmic wax ester accumulation during biofilm-driven substrate assimilation at the alkane--water interface by Marinobacter hydrocarbonoclasticus SP17. Res Microbiol 159: 137–144. doi: 10.1016/j.resmic.2007.11.013
|
| [9] | Baldi F, Ivos̆ević N, Minacci A, et al. (1999) Adhesion of Acinetobacter Venetianus to Diesel Fuel Droplets Studied with In Situ Electrochemical and Molecular Probes. Appl Environ Microbiol 65: 2041–2048. |
| [10] |
Macedo AJ, Kuhlicke U, Neu TR, et al. (2005) Three stages of a biofilm community developing at the liquid-liquid interface between polychlorinated biphenyls and water. Appl Environ Microbiol 71: 7301–7309. doi: 10.1128/AEM.71.11.7301-7309.2005
|
| [11] |
Pepi M, Minacci A, Di Cello F, et al. (2003) Long-term analysis of diesel fuel consumption in a co-culture of Acinetobacter Venetianus, Pseudomonas putida and Alcaligenes faecalis. Antonie Van Leeuwenhoek 83: 3–9. doi: 10.1023/A:1022930421705
|
| [12] |
Trejo M, Douarche C, Bailleux V, et al. (2013) Elasticity and wrinkled morphology of Bacillus subtilis pellicles. Proc Natl Acad Sci U S A 110: 2011–2016 doi: 10.1073/pnas.1217178110
|
| [13] |
Armitano J, Méjean V, Jourlin-Castelli C (2014) Gram-negative bacteria can also form pellicles. Environ Microbiol Rep 6: 534–544. doi: 10.1111/1758-2229.12171
|
| [14] |
Friedman L, Kolter R (2004) Two genetic loci produce distinct carbohydrate-rich structural components of the Pseudomonas aeruginosa biofilm matrix. J Bacteriol 186: 4457–4465. doi: 10.1128/JB.186.14.4457-4465.2004
|
| [15] |
Koza A, Hallett PD, Moon CD, et al. (2009) Characterization of a novel air-liquid interface biofilm of Pseudomonas fluorescens SBW25. Microbiol Read Engl 155: 1397–1406. doi: 10.1099/mic.0.025064-0
|
| [16] |
Sochorová Z, Petráčková D, Sitařová B, et al. (2014) Morphological and proteomic analysis of early stage air-liquid interface biofilm formation in Mycobacterium smegmatis. Microbiol Read Engl 160: 1346–1356. doi: 10.1099/mic.0.076174-0
|
| [17] | Lembre P, Lorentz C, Di Martino P (2012) Exopolysaccharides of the Biofilm Matrix: A Complex Biophysical World. In: Dr. Desiree Nedra Karunaratne Ed, The Complex World of Polysaccharides. InTech. |
| [18] |
Stewart PS, Rani SA, Gjersing E, et al. (2007) Observations of cell cluster hollowing in Staphylococcus epidermidis biofilms. Lett Appl Microbiol 44: 454–457. doi: 10.1111/j.1472-765X.2007.02112.x
|
| [19] |
Chen M-Y, Lee D-J, Tay J-H, et al. (2007) Staining of extracellular polymeric substances and cells in bioaggregates. Appl Microbiol Biotechnol 75: 467–474. doi: 10.1007/s00253-006-0816-5
|
| [20] |
Nosyk O, ter Haseborg E, Metzger U, et al. (2008) A standardized pre-treatment method of biofilm flocs for fluorescence microscopic characterization. J Microbiol Methods 75: 449–456. doi: 10.1016/j.mimet.2008.07.024
|
| [21] |
Wagner M, Ivleva NP, Haisch C, et al. (2009) Combined use of confocal laser scanning microscopy (CLSM) and Raman microscopy (RM): investigations on EPS-Matrix. Water Res 43: 63–76. doi: 10.1016/j.watres.2008.10.034
|
| [22] |
Sun C, Fiksdal L, Hanssen-Bauer A, et al. (2011) Characterization of membrane biofouling at different operating conditions (flux) in drinking water treatment using confocal laser scanning microscopy (CLSM) and image analysis. J Membr Sci 382: 194–201. doi: 10.1016/j.memsci.2011.08.010
|
| [23] | Murray JM (2005) Confocal microscopy, deconvolution, and structured illumination methods. In: Spector DL, Goldman RD, Basic methods in microscopy. Cold Spring Harbour Laboratory Press, New York. |
| [24] |
Baird FJ, Wadsworth MP, Hill JE (2012) Evaluation and optimization of multiple fluorophore analysis of a Pseudomonas aeruginosa biofilm. J Microbiol Methods 90: 192–196. doi: 10.1016/j.mimet.2012.05.004
|
| [25] |
Leriche V, Sibille P, Carpentier B (2000) Use of an enzyme-linked lectinsorbent assay to monitor the shift in polysaccharide composition in bacterial biofilms. Appl Environ Microbiol 66: 1851–1856. doi: 10.1128/AEM.66.5.1851-1856.2000
|
| [26] |
Zippel B, Neu TR (2011) Characterization of glycoconjugates of extracellular polymeric substances in tufa-associated biofilms by using fluorescence lectin-binding analysis. Appl Environ Microbiol 77: 505–516. doi: 10.1128/AEM.01660-10
|
| [27] | Decho AW, Kawaguchi T (1999) Confocal imaging of in situ natural microbial communities and their extracellular polymeric secretions using Nanoplast resin. Biotechnology 27: 1246–1252. |
| [28] |
Neu T, Swerhone GD, Lawrence JR (2001) Assessment of lectin-binding analysis for in situ detection of glycoconjugates in biofilm systems. Microbiol Read Engl 147: 299–313. doi: 10.1099/00221287-147-2-299
|
| [29] |
Neu TR, Kuhlicke U, Lawrence JR (2002) Assessment of fluorochromes for two-photon laser scanning microscopy of biofilms. Appl Environ Microbiol 68: 901–909. doi: 10.1128/AEM.68.2.901-909.2002
|
| [30] |
Yang Y, Sreenivasan PK, Subramanyam R, et al. (2006) Multiparameter assessments to determine the effects of sugars and antimicrobials on a polymicrobial oral biofilm. Appl Environ Microbiol 72: 6734–6742. doi: 10.1128/AEM.01013-06
|
| [31] |
Wigglesworth-Cooksey B, Cooksey KE (2005) Use of fluorophore-conjugated lectins to study cell-cell interactions in model marine biofilms. Appl Environ Microbiol 71: 428–435. doi: 10.1128/AEM.71.1.428-435.2005
|
| [32] |
Lawrence JR, Chenier MR, Roy R, et al. (2004) Microscale and molecular assessment of impacts of nickel, nutrients, and oxygen level on structure and function of river biofilm communities. Appl Environ Microbiol 70: 4326–4339. doi: 10.1128/AEM.70.7.4326-4339.2004
|
| [33] |
Villacorte LO, Ekowati Y, Neu TR, et al. (2015) Characterisation of algal organic matter produced by bloom-forming marine and freshwater algae. Water Res 73: 216–230. doi: 10.1016/j.watres.2015.01.028
|
| [34] |
Strathmann M, Wingender J, Flemming H-C (2002) Application of fluorescently labelled lectins for the visualization and biochemical characterization of polysaccharides in biofilms of Pseudomonas aeruginosa. J Microbiol Methods 50: 237–248. doi: 10.1016/S0167-7012(02)00032-5
|
| [35] |
Zhang RY, Neu TR, Bellenberg S, et al. (2015) Use of lectins to in situ visualize glycoconjugates of extracellular polymeric substances in acidophilic archaeal biofilms. Microb Biotechnol 8: 448–461. doi: 10.1111/1751-7915.12188
|
| [36] |
Johnsen AR, Hausner M, Schnell A, et al. (2000) Evaluation of fluorescently labeled lectins for noninvasive localization of extracellular polymeric substances in Sphingomonas biofilms. Appl Environ Microbiol 66: 3487–3491. doi: 10.1128/AEM.66.8.3487-3491.2000
|
| [37] |
Wrede C, Heller C, Reitner J, et al. (2008) Correlative light/electron microscopy for the investigation of microbial mats from Black Sea Cold Seeps. J Microbiol Methods 73: 85–91. doi: 10.1016/j.mimet.2008.02.020
|
| [38] |
Herzberg M, Elimelech M (2007) Biofouling of reverse osmosis membranes: Role of biofilm-enhanced osmotic pressure. J Membr Sci 295: 11–20. doi: 10.1016/j.memsci.2007.02.024
|
| [39] | Di Martino P, Doumèche B, Galas L, et al. (2007) Assessing chemical cleaning of nanofiltration membranes in a drinking water production plant: a combination of chemical composition analysis and fluorescence microscopy. Water Sci Technol 55: 219–225. |
| [40] |
Doumèche B, Galas L, Vaudry H, et al. (2007) Membrane Foulants Characterization in a Drinking Water Production Unit. Food Bioprod Process 85: 42–48. doi: 10.1205/fbp06020
|
| [41] | Tseng BS, Majerczyk CD, Passos da Silva D, et al. (2016) Quorum sensing influences Burkholderia thailandensis biofilm development and matrix production. J Bacteriol in press. |
| [42] |
Turonova H, Neu TR, Ulbrich P, et al. (2016) The biofilm matrix of Campylobacter jejuni determined by fluorescence lectin-binding analysis. Biofouling 32: 597–608. doi: 10.1080/08927014.2016.1169402
|
| [43] |
Bennke CM, Neu TR, Fuchs BM, et al. (2013) Mapping glycoconjugate-mediated interactions of marine Bacteroidetes with diatoms. Syst Appl Microbiol 36: 417–425. doi: 10.1016/j.syapm.2013.05.002
|
| [44] |
Peltola M, Neu TR, Raulio M, et al. (2008) Architecture of Deinococcus geothermalis biofilms on glass and steel: a lectin study. Environ Microbiol 10: 1752–1759. doi: 10.1111/j.1462-2920.2008.01596.x
|
| [45] |
Neu TR, Woelfl S, Lawrence JR (2004) Three-dimensional differentiation of photo-autotrophic biofilm constituents by multi-channel laser scanning microscopy (single-photon and two-photon excitation). J Microbiol Methods 56: 161–172. doi: 10.1016/j.mimet.2003.10.012
|
| [46] |
Houari A, Picard J, Habarou H, et al. (2008) Rheology of biofilms formed at the surface of NF membranes in a drinking water production unit. Biofouling 24: 235–240. doi: 10.1080/08927010802023764
|
| [47] | Harabi A, Bouzerara F (2011) Fabrication of Tubular Membrane Supports from Low Price Raw Materials, Using Both Centrifugal Casting and/or Extrusion Methods. INTECH Open Access Publisher, Chapter 13: 253-274 |
| [48] |
Böckelmann U, Manz W, Neu TR, et al. (2002) Investigation of lotic microbial aggregates by a combined technique of fluorescent in situ hybridization and lectin-binding-analysis. J Microbiol Methods 49: 75–87. doi: 10.1016/S0167-7012(01)00354-2
|
| [49] |
Houari A, Seyer D, Kecili K, et al. (2013) Kinetic development of biofilm on NF membranes at the Méry-sur-Oise plant, France. Biofouling 29: 109–118. doi: 10.1080/08927014.2012.752464
|
| [50] |
Marconnet C, Houari A, Galas L, et al. (2009) Biodegradable dissolved organic carbon concentration of feed water and NF membrane biofouling: a pilot train study. Desalination 242: 228–235. doi: 10.1016/j.desal.2008.04.009
|
| [51] |
Laue H, Schenk A, Li H, et al. (2006) Contribution of alginate and levan production to biofilm formation by Pseudomonas syringae. Microbiol Read Engl 152: 2909–2918. doi: 10.1099/mic.0.28875-0
|
| [52] |
Jennings LK, Storek KM, Ledvina HE, et al. (2015) Pel is a cationic exopolysaccharide that cross-links extracellular DNA in the Pseudomonas aeruginosa biofilm matrix. Proc Natl Acad Sci U S A 112: 11353–11358. doi: 10.1073/pnas.1503058112
|
| [53] |
Staudt C, Horn H, Hempel DC, et al. (2004) Volumetric measurements of bacterial cells and extracellular polymeric substance glycoconjugates in biofilms. Biotechnol Bioeng 88: 585–592. doi: 10.1002/bit.20241
|
| [54] |
Sanford BA, Thomas VL, Mattingly SJ, et al. (1995) Lectin-biotin assay for slime present in in situ biofilm produced by Staphylococcus epidermidis using transmission electron microscopy (TEM). J Ind Microbiol 15: 156–161. doi: 10.1007/BF01569820
|
| [55] | Maeyama R, Mizunoe Y, Anderson JM, et al. (2004) Confocal imaging of biofilm formation process using fluoroprobed Escherichia coli and fluoro-stained exopolysaccharide. J Biomed Mater Res A 70: 274–282. |
| [56] |
Bahulikar RA, Kroth PG (2007) Localization of EPS components secreted by freshwater diatoms using differential staining with fluorophore-conjugated lectins and other fluorochromes. Eur J Phycol 42: 199–208. doi: 10.1080/09670260701289779
|
| [57] |
Hao L, Guo Y, Byrne JM, et al. (2016) Binding of heavy metal ions in aggregates of microbial cells, EPS and biogenic iron minerals measured in-situ using metal- and glycoconjugates-specific fluorophores. Geochim Cosmochim Acta 180: 66–96. doi: 10.1016/j.gca.2016.02.016
|
| [58] |
Lawrence JR, Swerhone GDW, Kuhlicke U, et al. (2007) In situ evidence for microdomains in the polymer matrix of bacterial microcolonies. Can J Microbiol 53: 450–458. doi: 10.1139/W06-146
|
| [59] | WHO (2011) Guidelines for drinking-water quality, 4 ed. World Health Organization, Geneva. |
| [60] |
Cyna B, Chagneau G, Bablon G, et al. (2002) Two years of nanofiltration at the Méry-sur-Oise plant, France. Desalination 147: 69–75. doi: 10.1016/S0011-9164(02)00578-7
|
| [61] |
Hilal N, Al-Zoubi H, Darwish NA, et al. (2004) A comprehensive review of nanofiltration membranes: Treatment, pretreatment, modelling, and atomic force microscopy. Desalination 170: 281–308. doi: 10.1016/j.desal.2004.01.007
|
| [62] |
Speth TF, Summers RS, Gusses AM (1998) Nanofiltration Foulants from a Treated Surface Water. Environ Sci Technol 32: 3612–3617. doi: 10.1021/es9800434
|
| [63] |
Vrouwenvelder HS, van Paassen JAM, Folmer HC, et al. (1998) Biofouling of membranes for drinking water production. Desalination 118: 157–166. doi: 10.1016/S0011-9164(98)00116-7
|
| [64] |
Liikanen R, Yli-Kuivila J, Laukkanen R (2002) Efficiency of various chemical cleanings for nanofiltration membrane fouled by conventionally-treated surface water. J Membr Sci 195: 265–276. doi: 10.1016/S0376-7388(01)00569-5
|
| [65] |
Lappin‐Scott HM, Costerton JW (1989) Bacterial biofilms and surface fouling. Biofouling 1: 323–342. doi: 10.1080/08927018909378120
|
| [66] |
Flemming H-C, Schaule G, Griebe T, et al. (1997) Biofouling—the Achilles heel of membrane processes. Desalination 113: 215–225. doi: 10.1016/S0011-9164(97)00132-X
|
| [67] |
Violleau D, Essis-Tome H, Habarou H, et al. (2005) Fouling studies of a polyamide nanofiltration membrane by selected natural organic matter: an analytical approach. Desalination 173: 223–238. doi: 10.1016/j.desal.2004.07.048
|
| [68] |
Koyuncu I, Wiesner MR, Bele C, et al. (2006) Bench-scale assessment of pretreatment to reduce fouling of salt-rejecting membranes. Desalination 197: 94–105. doi: 10.1016/j.desal.2005.10.041
|
| [69] |
Shadpour H, Musyimi H, Chen J, et al. (2006) Physiochemical properties of various polymer substrates and their effects on microchip electrophoresis performance. J Chromatogr A 1111: 238–251. doi: 10.1016/j.chroma.2005.08.083
|
| [70] |
Ivnitsky H, Katz I, Minz D, et al. (2007) Bacterial community composition and structure of biofilms developing on nanofiltration membranes applied to wastewater treatment. Water Res 41: 3924–3935. doi: 10.1016/j.watres.2007.05.021
|
| [71] |
Derlon N, Grütter A, Brandenberger F, et al. (2016) The composition and compression of biofilms developed on ultrafiltration membranes determine hydraulic biofilm resistance. Water Res 102: 63-72 doi: 10.1016/j.watres.2016.06.019
|
| [72] |
Vrouwenvelder JS, Manolarakis SA, van der Hoek JP, et al. (2008) Quantitative biofouling diagnosis in full scale nanofiltration and reverse osmosis installations. Water Res 42: 4856–4868. doi: 10.1016/j.watres.2008.09.002
|
| [73] | Ridgway H, Flemming HC (1996) Membrane Biofouling, In: Water Treatment Membrane Processes. McGraw Hill, New York. |
| [74] |
Speth TF, Gusses AM, Scott Summers R (2000) Evaluation of nanofiltration pretreatments for flux loss control. Desalination 130: 31–44. doi: 10.1016/S0011-9164(00)00072-2
|
| [75] |
Marconnet C, Houari A, Seyer D, et al. (2011) Membrane biofouling control by UV irradiation. Desalination 276: 75–81. doi: 10.1016/j.desal.2011.03.016
|
| [76] |
Li Q, Elimelech M (2004) Organic fouling and chemical cleaning of nanofiltration membranes: measurements and mechanisms. Environ Sci Technol 38: 4683–4693. doi: 10.1021/es0354162
|
| [77] |
Houari A, Seyer D, Couquard F, et al. (2010) Characterization of the biofouling and cleaning efficiency of nanofiltration membranes. Biofouling 26: 15–21. doi: 10.1080/08927010903277749
|
| [78] |
Zhu H, Nyström M (1998) Cleaning results characterized by flux, streaming potential and FTIR measurements. Colloids Surf Physicochem Eng Asp 138: 309–321. doi: 10.1016/S0927-7757(97)00072-1
|

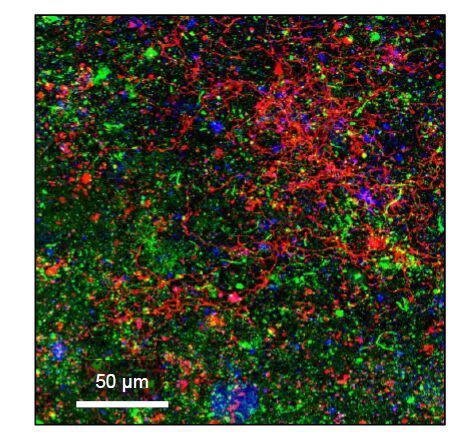

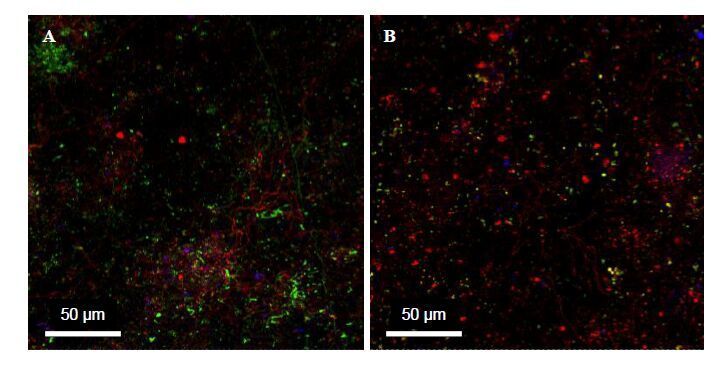
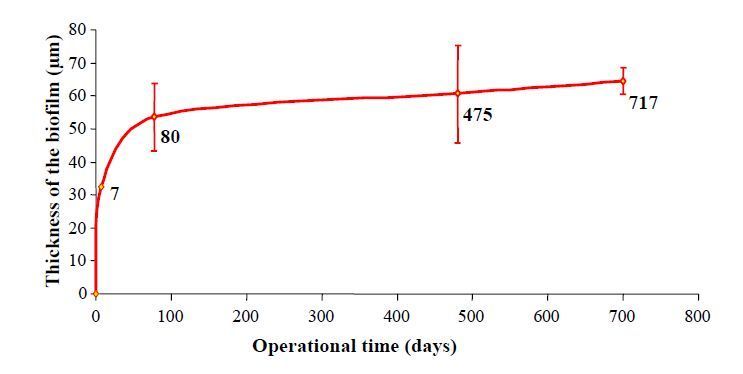
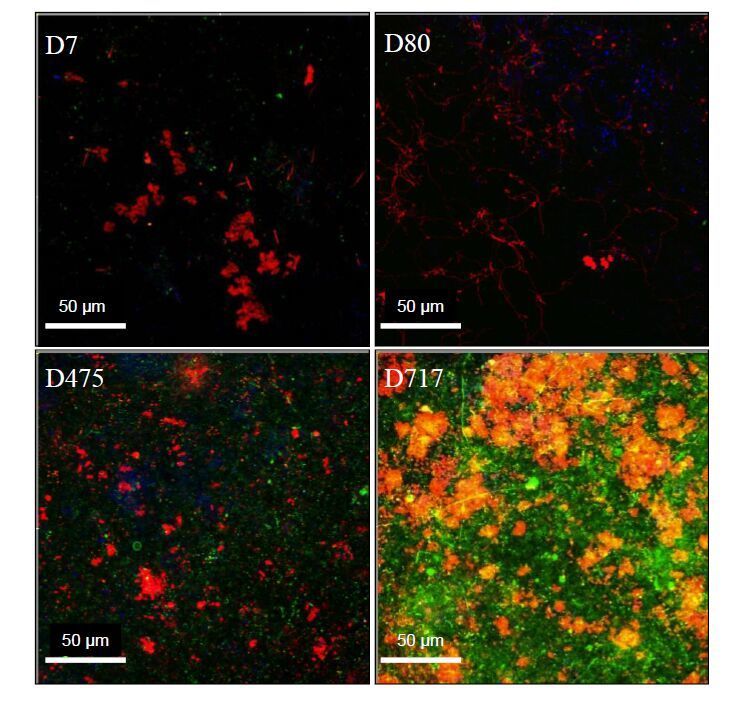
Figures(8) / Tables(1)

Mohamed Amine Ben Mlouka, Thomas Cousseau, Patrick Di Martino. Application of fluorescently labelled lectins for the study of polysaccharides in biofilms with a focus on biofouling of nanofiltration membranes[J]. AIMS Molecular Science, 2016, 3(3): 338-356. doi: 10.3934/molsci.2016.3.338







 DownLoad:
DownLoad: