We propose a numerical-based approach extending the conditional MVAR Granger causality (MVGC) analysis for the construction of directed connectivity networks in the presence of both exogenous/stimuli and modulatory inputs. The performance of the proposed scheme is validated using both synthetic stochastic data considering also the influence of haemodynamics latencies and a benchmark fMRI dataset related to the role of attention in the perception of visual motion. The particular fMRI dataset has been used in many studies to evaluate alternative model hypotheses using the Dynamic Causal Modelling (DCM) approach. Based on the use of the Bayes factor, we show that the obtained GC connectivity network compares well to a reference model that has been selected through DCM analysis among other candidate models. Thus, our findings suggest that the proposed scheme can be successfully used as a stand-alone or complementary to DCM approach to find directed causal connectivity patterns in task-related fMRI studies.
Citation: Evangelos Almpanis, Constantinos Siettos. Construction of functional brain connectivity networks from fMRI data with driving and modulatory inputs: an extended conditional Granger causality approach[J]. AIMS Neuroscience, 2020, 7(2): 66-88. doi: 10.3934/Neuroscience.2020005
We propose a numerical-based approach extending the conditional MVAR Granger causality (MVGC) analysis for the construction of directed connectivity networks in the presence of both exogenous/stimuli and modulatory inputs. The performance of the proposed scheme is validated using both synthetic stochastic data considering also the influence of haemodynamics latencies and a benchmark fMRI dataset related to the role of attention in the perception of visual motion. The particular fMRI dataset has been used in many studies to evaluate alternative model hypotheses using the Dynamic Causal Modelling (DCM) approach. Based on the use of the Bayes factor, we show that the obtained GC connectivity network compares well to a reference model that has been selected through DCM analysis among other candidate models. Thus, our findings suggest that the proposed scheme can be successfully used as a stand-alone or complementary to DCM approach to find directed causal connectivity patterns in task-related fMRI studies.

| [1] |
Friston KJ (1994) Functional and effective connectivity in neuroimaging: A synthesis. Hum Brain Mapp 2: 56-78. doi: 10.1002/hbm.460020107

|
| [2] |
Friston KJ (2011) Functional and effective connectivity: A review. Brain Connect 1: 13-36. doi: 10.1089/brain.2011.0008
|
| [3] |
Friston K, Moran R, Seth AK (2013) Analysing connectivity with granger causality and dynamic causal modelling. Curr Opin Neurobiol 23: 172-178. doi: 10.1016/j.conb.2012.11.010
|
| [4] |
Siettos C, Starke J (2016) Multiscale modeling of brain dynamics: from single neurons and networks to mathematical tools. Wiley Interdiscip Rev Syst Biol Med 8: 438-458. doi: 10.1002/wsbm.1348
|
| [5] |
Smith SM, Miller KL, Salimi-Khorshidi G, et al. (2011) Network modelling methods for fmri. Neuroimage 54: 875-891. doi: 10.1016/j.neuroimage.2010.08.063
|
| [6] |
Brazier MA (1972) Spread of seizure discharges in epilepsy: Anatomical and electrophysiological considerations. Exp Neurol 36: 263-272. doi: 10.1016/0014-4886(72)90022-2
|
| [7] |
Gerstein GL, Perkel DH, Subramanian K (1978) Identification of functionally related neural assemblies. Brain Res 140: 43-62. doi: 10.1016/0006-8993(78)90237-8
|
| [8] |
Zalesky A, Fornito A, Bullmore E (2012) On the use of correlation as a measure of network connectivity. NeuroImage 60: 2096-2106. doi: 10.1016/j.neuroimage.2012.02.001
|
| [9] |
Siggiridou E, Kugiumtzis D, Kimiskidis V (2014) Correlation networks for identifying changes in brain connectivity during epileptiform discharges and transcranial magnetic stimulation. Sensors 14: 12585-12597. doi: 10.3390/s140712585
|
| [10] | Hockett CF (1953) The mathematical theory of communication. |
| [11] |
Vlachos I, Kugiumtzis D (2010) Nonuniform state-space reconstruction and coupling detection. Phys Rev E 82: 016207. doi: 10.1103/PhysRevE.82.016207
|
| [12] |
Vicente R, Wibral M, Lindner M, et al. (2011) Transfer entropy—a model-free measure of effective connectivity for the neurosciences. J Comput Neurosci 30: 45-67. doi: 10.1007/s10827-010-0262-3
|
| [13] |
Savva AD, Mitsis GD, Matsopoulos GK (2019) Assessment of dynamic functional connectivity in resting-state fMRI using the sliding window technique. Brain Behav 9: e01255. doi: 10.1002/brb3.1255
|
| [14] |
Lachaux JP, Rodriguez E, Martinerie J, et al. (1999) Measuring phase synchrony in brain signals. Hum Brain Mapp 8: 194-208. doi: 10.1002/(SICI)1097-0193(1999)8:4<194::AID-HBM4>3.0.CO;2-C
|
| [15] |
Stam CJ, Nolte G, Daffertshofer A (2007) Phase lag index: Assessment of functional connectivity from multi channel EEG and MEG with diminished bias from common sources. Hum Brain Mapp 28: 1178-1193. doi: 10.1002/hbm.20346
|
| [16] |
Glerean E, Salmi J, Lahnakoski JM, et al. (2012) Functional magnetic resonance imaging phase synchronization as a measure of dynamic functional connectivity. Brain Connect 2: 91-101. doi: 10.1089/brain.2011.0068
|
| [17] |
Mylonas DS, Siettos CI, Evdokimidis I, et al. (2015) Modular patterns of phase desynchronization networks during a simple visuomotor task. Brain Topogr 29: 118-129. doi: 10.1007/s10548-015-0451-5
|
| [18] |
Calhoun VD, Adali T, Pearlson GD, et al. (2001) A method for making group inferences from functional mri data using independent component analysis. Human Brain Mapp 14: 140-151. doi: 10.1002/hbm.1048
|
| [19] |
Calhoun VD, Liu J, Adali T (2009) A review of group ica for fmri data and ica for joint inference of imaging, genetic, and erp data. Neuroimage 45: S163-S172. doi: 10.1016/j.neuroimage.2008.10.057
|
| [20] |
Reidl J, Starke J, Omer DB, et al. (2007) Independent component analysis of high-resolution imaging data identifies distinct functional domains. Neuroimage 34: 94-108. doi: 10.1016/j.neuroimage.2006.08.031
|
| [21] | Anderson A, Cohen MS (2013) Decreased small-world functional network connectivity and clustering across resting state networks in schizophrenia: an fmri classification tutorial. Front Hum Neurosci 7: 520. |
| [22] |
Tenenbaum JB, De Silva V, Langford JC (2000) A global geometric framework for nonlinear dimensionality reduction. Science 290: 2319-2323. doi: 10.1126/science.290.5500.2319
|
| [23] |
Duncan D, Talmon R, Zaveri HP, et al. (2013) Identifying preseizure state in intracranial EEG data using diffusion kernels. Math Biosci Eng 10: 579-590. doi: 10.3934/mbe.2013.10.579
|
| [24] |
Granger CW (1969) Investigating causal relations by econometric models and cross-spectral methods. Econometrica 424-438. doi: 10.2307/1912791
|
| [25] |
Granger C, Newbold P (1974) Spurious regressions in econometrics. J Econom 2: 111-120. doi: 10.1016/0304-4076(74)90034-7
|
| [26] |
Geweke J (1982) Measurement of linear dependence and feedback between multiple time series. J Am Stat Assoc 77: 304-313. doi: 10.1080/01621459.1982.10477803
|
| [27] |
Seth AK (2010) A matlab toolbox for granger causal connectivity analysis. J Neurosci Methods 186: 262-273. doi: 10.1016/j.jneumeth.2009.11.020
|
| [28] |
Barnett L, Seth AK (2014) The mvgc multivariate granger causality toolbox: A new approach to granger-causal inference. J Neurosci Methods 223: 50-68. doi: 10.1016/j.jneumeth.2013.10.018
|
| [29] |
Seth AK, Barrett AB, Barnett L (2015) Granger causality analysis in neuroscience and neuroimaging. J Neurosci 35: 3293-3297. doi: 10.1523/JNEUROSCI.4399-14.2015
|
| [30] |
Friston KJ, Harrison L, Penny W (2003) Dynamic causal modelling. Neuroimage 19: 1273-1302. doi: 10.1016/S1053-8119(03)00202-7
|
| [31] |
Keller CJ, Bickel S, Honey CJ, et al. (2013) Neurophysiological investigation of spontaneous correlated and anticorrelated fluctuations of the BOLD signal. J Neurosci 33: 6333-6342. doi: 10.1523/JNEUROSCI.4837-12.2013
|
| [32] |
Megumi F, Yamashita A, Kawato M, et al. (2015) Functional MRI neurofeedback training on connectivity between two regions induces long-lasting changes in intrinsic functional network. Front Hum Neurosci 9: 160. doi: 10.3389/fnhum.2015.00160
|
| [33] | Wiener N (1956) The theory of prediction. Modern Math Eng 1: 125-139. |
| [34] |
Barrett AB, Murphy M, Bruno MA, et al. (2012) Granger causality analysis of steady-state electroencephalographic signals during propofol-induced anaesthesia. PloS One 7: e29072. doi: 10.1371/journal.pone.0029072
|
| [35] |
Barnett L, Seth AK (2011) Behaviour of granger causality under filtering: theoretical invariance and practical application. J Neurosci Methods 201: 404-419. doi: 10.1016/j.jneumeth.2011.08.010
|
| [36] |
Gow DW, Segawa JA, Ahlfors SP, et al. (2008) Lexical influences on speech perception: a granger causality analysis of meg and eeg source estimates. Neuroimage 43: 614-623. doi: 10.1016/j.neuroimage.2008.07.027
|
| [37] |
Keil A, Sabatinelli D, Ding M, et al. (2009) Re-entrant projections modulate visual cortex in affective perception: Evidence from granger causality analysis. Hum Brain Mapp 30: 532-540. doi: 10.1002/hbm.20521
|
| [38] |
Nicolaou N, Hourris S, Alexandrou P, et al. (2012) Eeg-based automatic classification of ‘awake’ versus ‘anesthetized’ state in general anesthesia using granger causality. PloS One 7: e33869. doi: 10.1371/journal.pone.0033869
|
| [39] |
De Tommaso M, Stramaglia S, Marinazzo D, et al. (2013) Functional and effective connectivity in eeg alpha and beta bands during intermittent flash stimulation in migraine with and without aura. Cephalalgia 33: 938-947. doi: 10.1177/0333102413477741
|
| [40] |
Protopapa F, Siettos CI, Evdokimidis I, et al. (2014) Granger causality analysis reveals distinct spatio-temporal connectivity patterns in motor and perceptual visuo-spatial working memory. Front Comput Neurosci 8: 146. doi: 10.3389/fncom.2014.00146
|
| [41] |
Protopapa F, Siettos CI, Myatchin I, et al. (2016) Children with well controlled epilepsy possess different spatio-temporal patterns of causal network connectivity during a visual working memory task. Cogn Neurodyn 10: 99-111. doi: 10.1007/s11571-015-9373-x
|
| [42] |
Al-Aidroos N, Said CP, Turk-Browne NB (2012) Top-down attention switches coupling between low-level and high-level areas of human visual cortex. Proc Natl Acad Sci 109: 14675-14680. doi: 10.1073/pnas.1202095109
|
| [43] |
Florin E, Gross J, Pfeifer J, et al. (2010) The effect of filtering on granger causality based multivariate causality measures. Neuroimage 50: 577-588. doi: 10.1016/j.neuroimage.2009.12.050
|
| [44] |
Zhou Z, Wang X, Klahr NJ, et al. (2011) A conditional granger causality model approach for group analysis in functional magnetic resonance imaging. Magn Reson Imaging 29: 418-433. doi: 10.1016/j.mri.2010.10.008
|
| [45] |
Wu G, Liao W, Stramaglia S, et al. (2013) Recovering directed networks in neuroimaging datasets using partially conditioned granger causality. BMC Neurosci 14: P260. doi: 10.1186/1471-2202-14-S1-P260
|
| [46] |
Tang W, Bressler SL, Sylvester CM, et al. (2012) Measuring granger causality between cortical regions from voxelwise fmri bold signals with lasso. PLoS Comput Biol 8: e1002513. doi: 10.1371/journal.pcbi.1002513
|
| [47] |
Roebroeck A, Formisano E, Goebel R (2005) Mapping directed influence over the brain using granger causality and fmri. Neuroimage 25: 230-242. doi: 10.1016/j.neuroimage.2004.11.017
|
| [48] |
Sabatinelli D, McTeague LM, Dhamala M, et al. (2015) Reduced medial prefrontal–subcortical connectivity in dysphoria: Granger causality analyses of rapid functional magnetic resonance imaging. Brain Connect 5: 1-9. doi: 10.1089/brain.2013.0186
|
| [49] |
Miao X, Wu X, Li R, et al. (2011) Altered connectivity pattern of hubs in default-mode network with alzheimer's disease: an granger causality modeling approach. PloS One 6: e25546. doi: 10.1371/journal.pone.0025546
|
| [50] |
Liao W, Ding J, Marinazzo D, et al. (2011) Small-world directed networks in the human brain: Multivariate granger causality analysis of resting-state fMRI. Neuroimage 54: 2683-2694. doi: 10.1016/j.neuroimage.2010.11.007
|
| [51] |
Gao Q, Duan X, Chen H (2011) Evaluation of effective connectivity of motor areas during motor imagery and execution using conditional granger causality. Neuroimage 54: 1280-1288. doi: 10.1016/j.neuroimage.2010.08.071
|
| [52] |
Lin FH, Ahveninen J, Raij T, et al. (2014) Increasing fMRI sampling rate improves granger causality estimates. PLoS One 9: e100319. doi: 10.1371/journal.pone.0100319
|
| [53] |
Duggento A, Passamonti L, Valenza G, et al. (2018) Multivariate granger causality unveils directed parietal to prefrontal cortex connectivity during task-free MRI. Sci Rep 8: 1-11. doi: 10.1038/s41598-018-23996-x
|
| [54] |
Penny WD, Stephan KE, Mechelli A, et al. (2004) Comparing dynamic causal models. Neuroimage 22: 1157-1172. doi: 10.1016/j.neuroimage.2004.03.026
|
| [55] |
Friston KJ, Bastos A, Litvak V, et al. (2012) Dcm for complex-valued data: cross-spectra, coherence and phase-delays. Neuroimage 59: 439-455. doi: 10.1016/j.neuroimage.2011.07.048
|
| [56] |
Guo S, Seth AK, Kendrick KM, et al. (2008) Partial granger causality—eliminating exogenous inputs and latent variables. J Neurosci Methods 172: 79-93. doi: 10.1016/j.jneumeth.2008.04.011
|
| [57] |
Roelstraete B, Rosseel Y (2012) Does partial granger causality really eliminate the influence of exogenous inputs and latent variables? J Neurosci Methods 206: 73-77. doi: 10.1016/j.jneumeth.2012.01.010
|
| [58] |
Ge T, Kendrick KM, Feng J (2009) A novel extended granger causal model approach demonstrates brain hemispheric differences during face recognition learning. PLoS Comput Biol 5: e1000570. doi: 10.1371/journal.pcbi.1000570
|
| [59] |
Bajaj S, Adhikari BM, Friston KJ, et al. (2016) Bridging the gap: Dynamic causal modeling and granger causality analysis of resting state functional magnetic resonance imaging. Brain Connect 6: 652-661. doi: 10.1089/brain.2016.0422
|
| [60] |
Dhamala M, Rangarajan G, Ding M (2008) Analyzing information flow in brain networks with nonparametric granger causality. Neuroimage 41: 354-362. doi: 10.1016/j.neuroimage.2008.02.020
|
| [61] |
Fox MD, Snyder AZ, Vincent JL, et al. (2005) The human brain is intrinsically organized into dynamic, anticorrelated functional networks. Proc Natl Acad Sci 102: 9673-9678. doi: 10.1073/pnas.0504136102
|
| [62] |
Biswal BB (2012) Resting state fMRI: A personal history. Neuroimage 62: 938-944. doi: 10.1016/j.neuroimage.2012.01.090
|
| [63] |
Seth AK, Chorley P, Barnett LC (2013) Granger causality analysis of fMRI BOLD signals is invariant to hemodynamic convolution but not downsampling. Neuroimage 65: 540-555. doi: 10.1016/j.neuroimage.2012.09.049
|
| [64] |
Ding M, Chen Y, Bressler SL (2006) Granger causality: basic theory and application to neuroscience. Handbook of time series analysis: recent theoretical developments and applications 437: 437-460. doi: 10.1002/9783527609970.ch17
|
| [65] |
Baccalá LA, Sameshima K (2001) Partial directed coherence: A new concept in neural structure determination. Biol Cybern 84: 463-474. doi: 10.1007/PL00007990
|
| [66] |
Dumoulin S, Wandell B (2008) Population receptive field estimates in human visual cortex. Neuroimage 39: 647. doi: 10.1016/j.neuroimage.2007.09.034
|
| [67] |
Büchel C, Friston K (1997) Modulation of connectivity in visual pathways by attention: cortical interactions evaluated with structural equation modelling and fmri. Cereb Cortex 7: 768-778. doi: 10.1093/cercor/7.8.768
|
| [68] |
Büchel C, Josephs O, Rees G, et al. (1998) The functional anatomy of attention to visual motion. a functional mri study. Brain 121: 1281-1294. doi: 10.1093/brain/121.7.1281
|
| [69] |
Penny WD, Stephan KE, Mechelli A, et al. (2004) Modelling functional integration: a comparison of structural equation and dynamic causal models. Neuroimage 23: S264-S274. doi: 10.1016/j.neuroimage.2004.07.041
|
| [70] | Penny WD, Friston KJ, Ashburner JT, et al. (2011) Statistical parametric mapping: the analysis of functional brain images Academic press. |
| [71] |
Friston KJ, Holmes AP, Worsley KJ, et al. (1994) Statistical parametric maps in functional imaging: a general linear approach. Human Brain Mapping 2: 189-210. doi: 10.1002/hbm.460020402
|
| [72] |
Worsley KJ, Friston KJ (1995) Analysis of fmri time-series revisited—again. Neuroimage 2: 173-181. doi: 10.1006/nimg.1995.1023
|
| [73] |
Zou C, Feng J (2009) Granger causality vs. dynamic bayesian network inference: A comparative study. BMC Bioinformatics 10: 122. doi: 10.1186/1471-2105-10-122
|
 neurosci-07-02-005-s001.txt neurosci-07-02-005-s001.txt |
 |
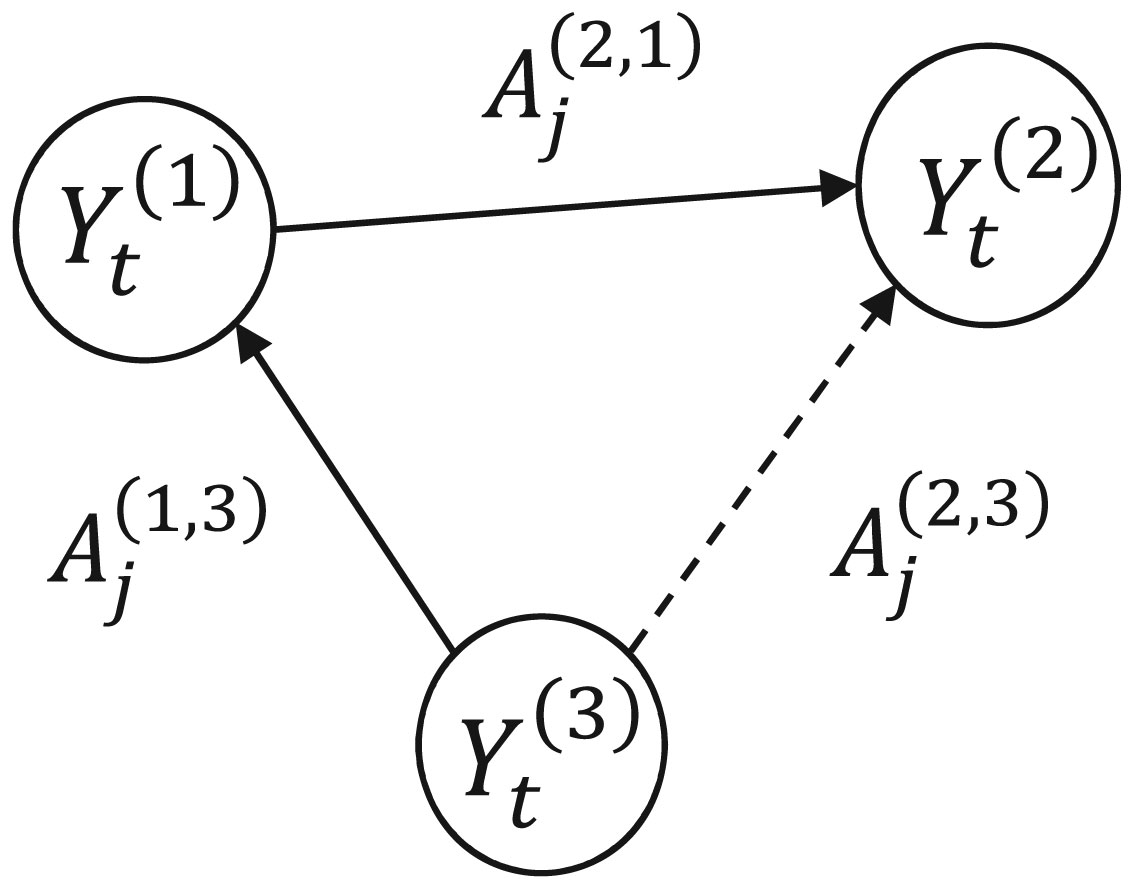
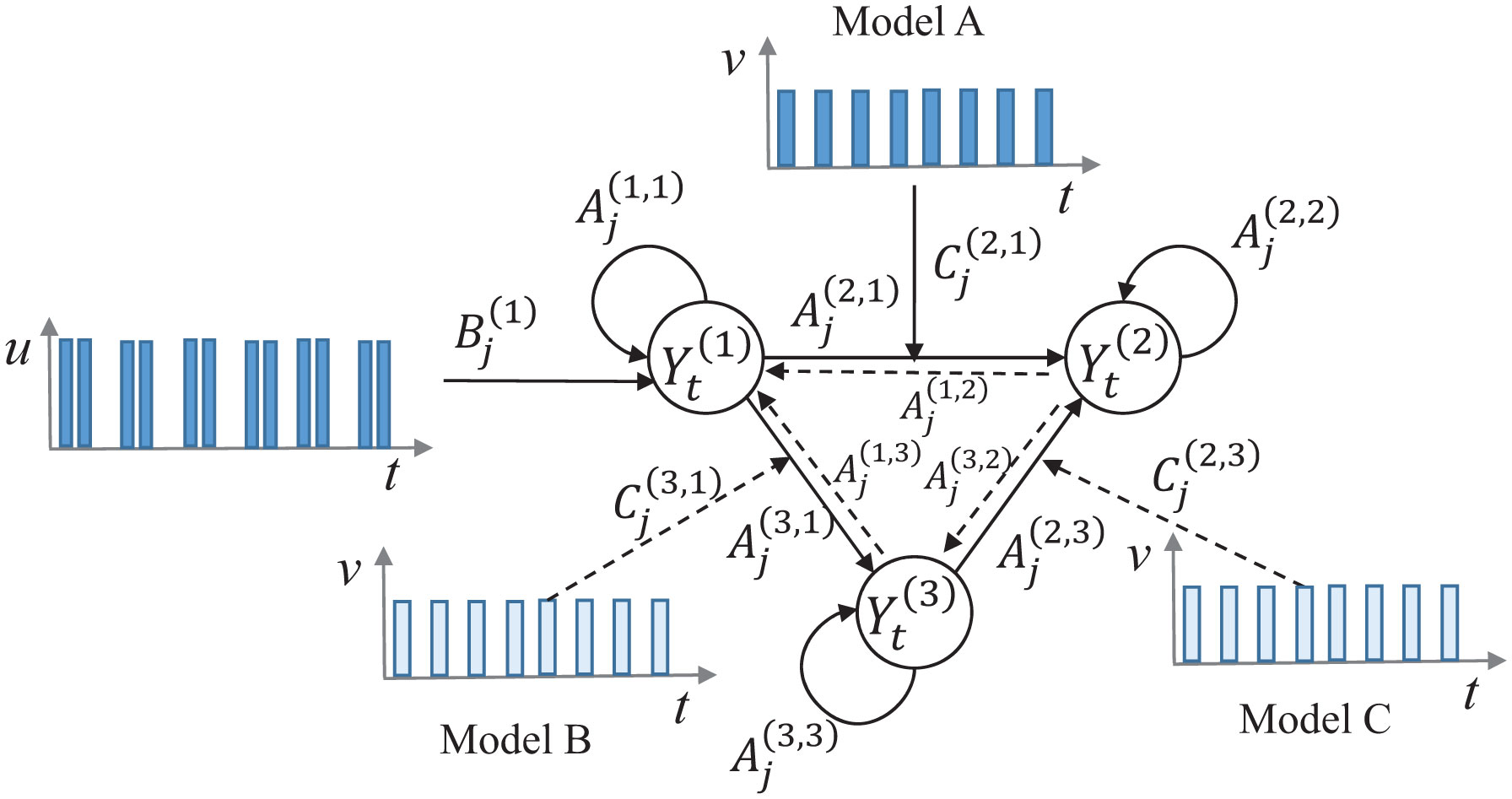
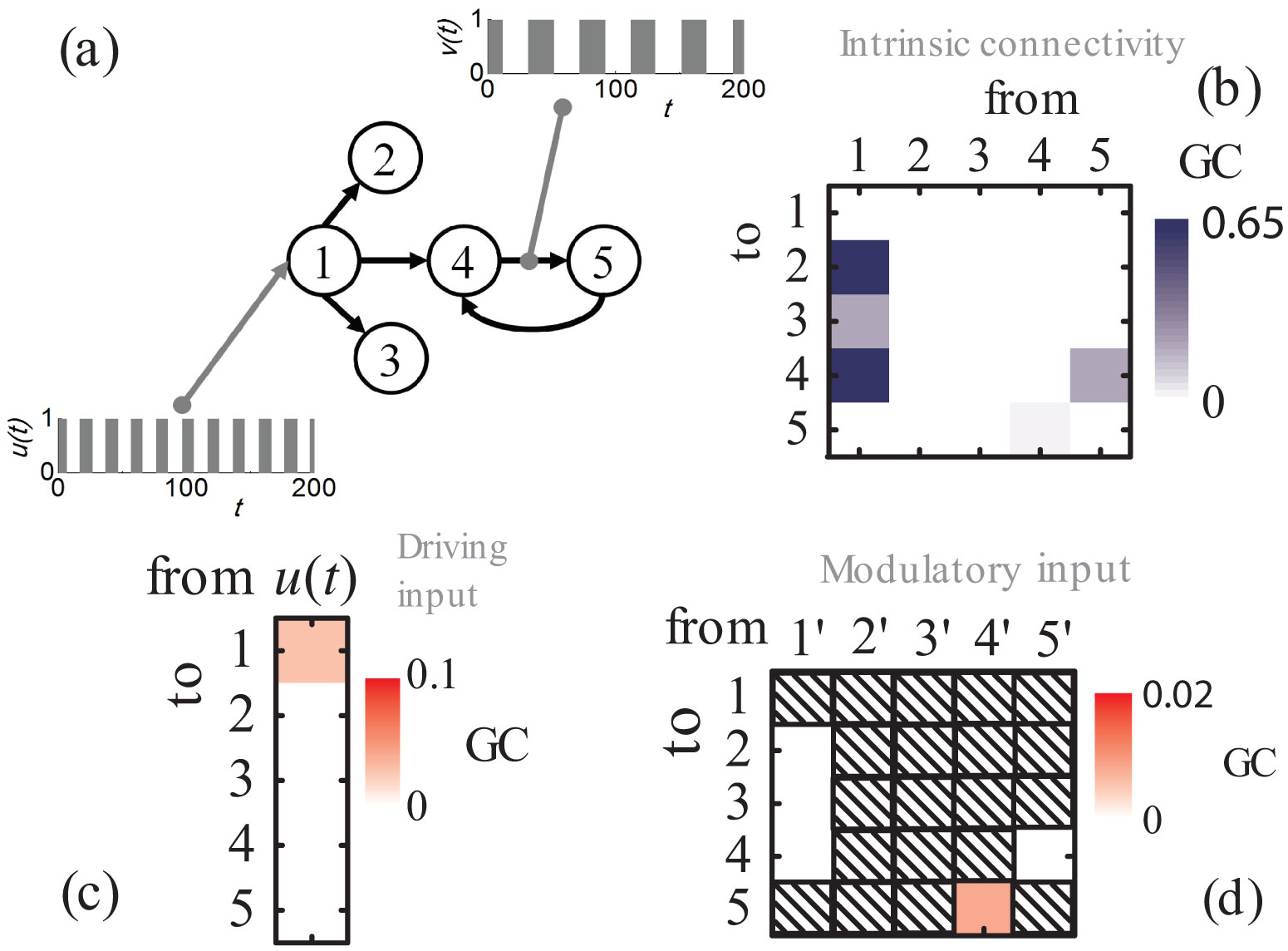
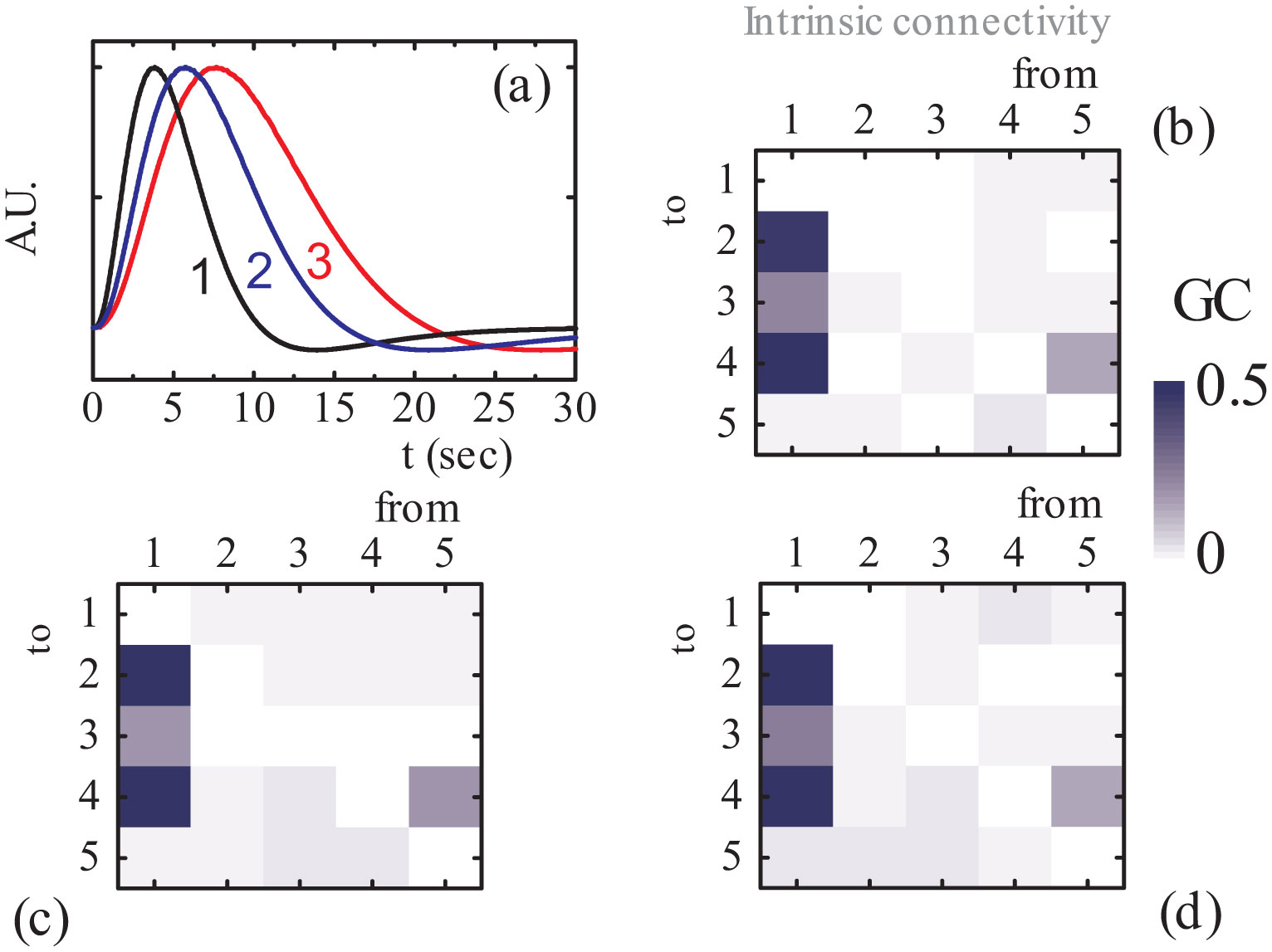
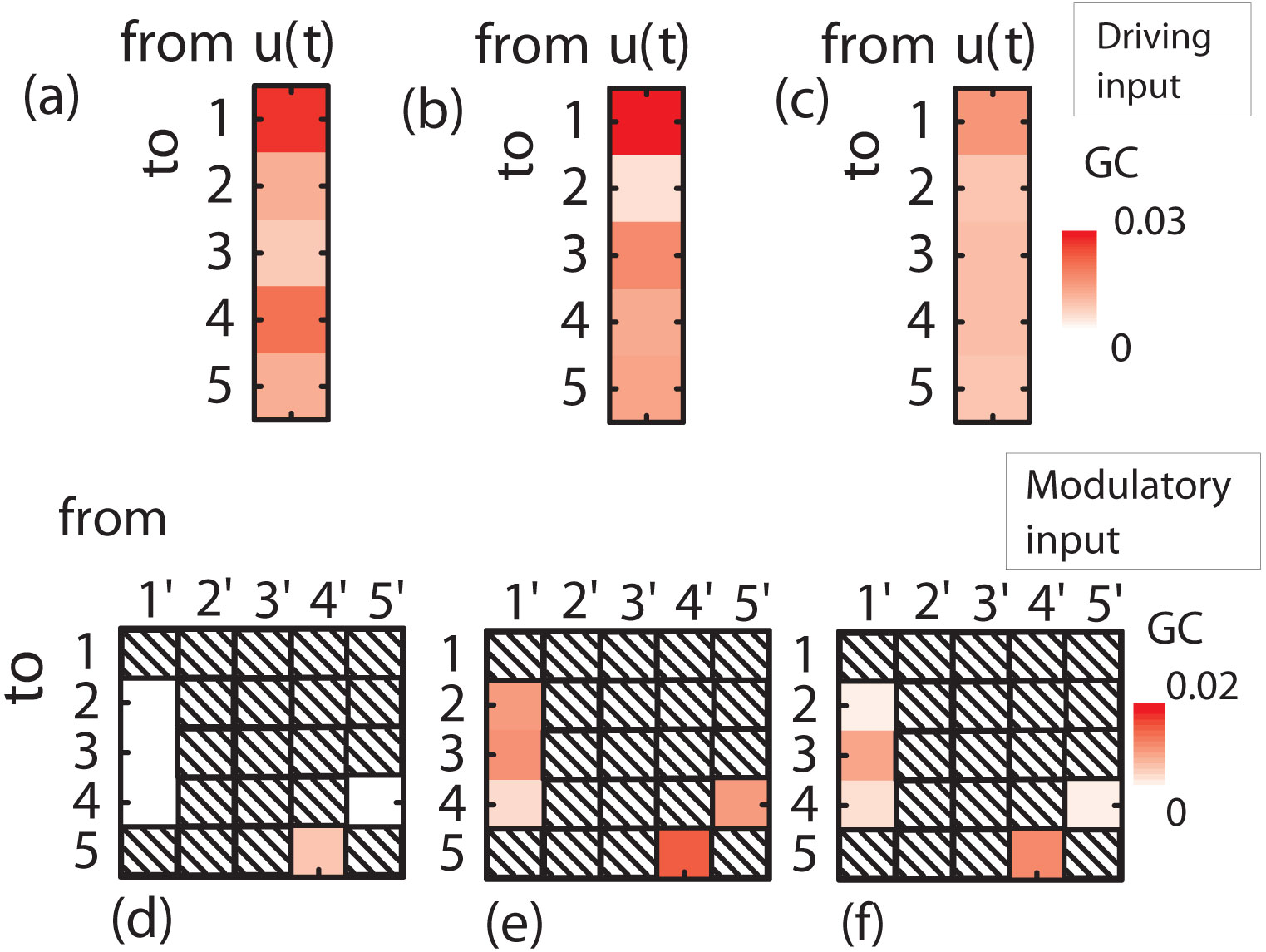
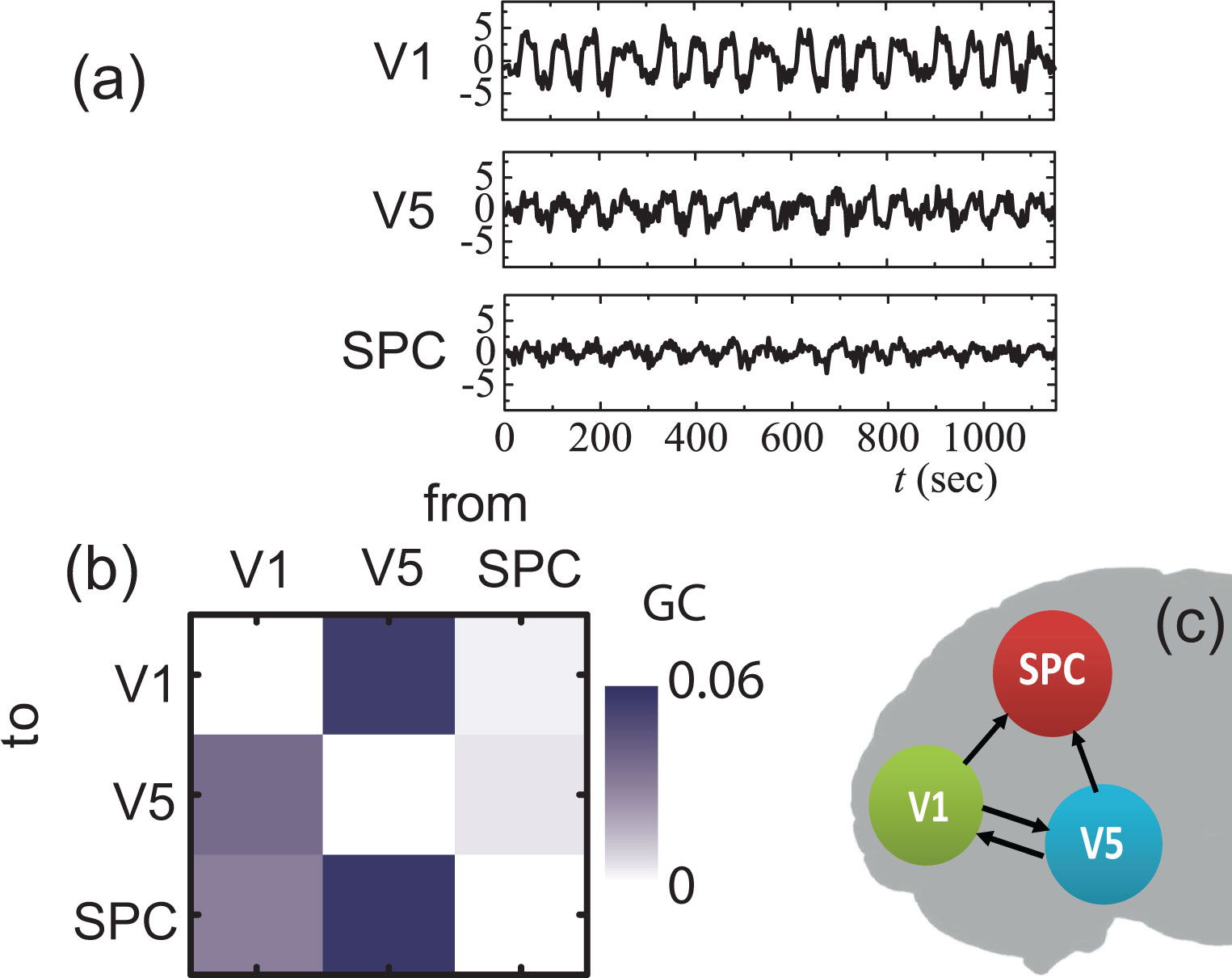
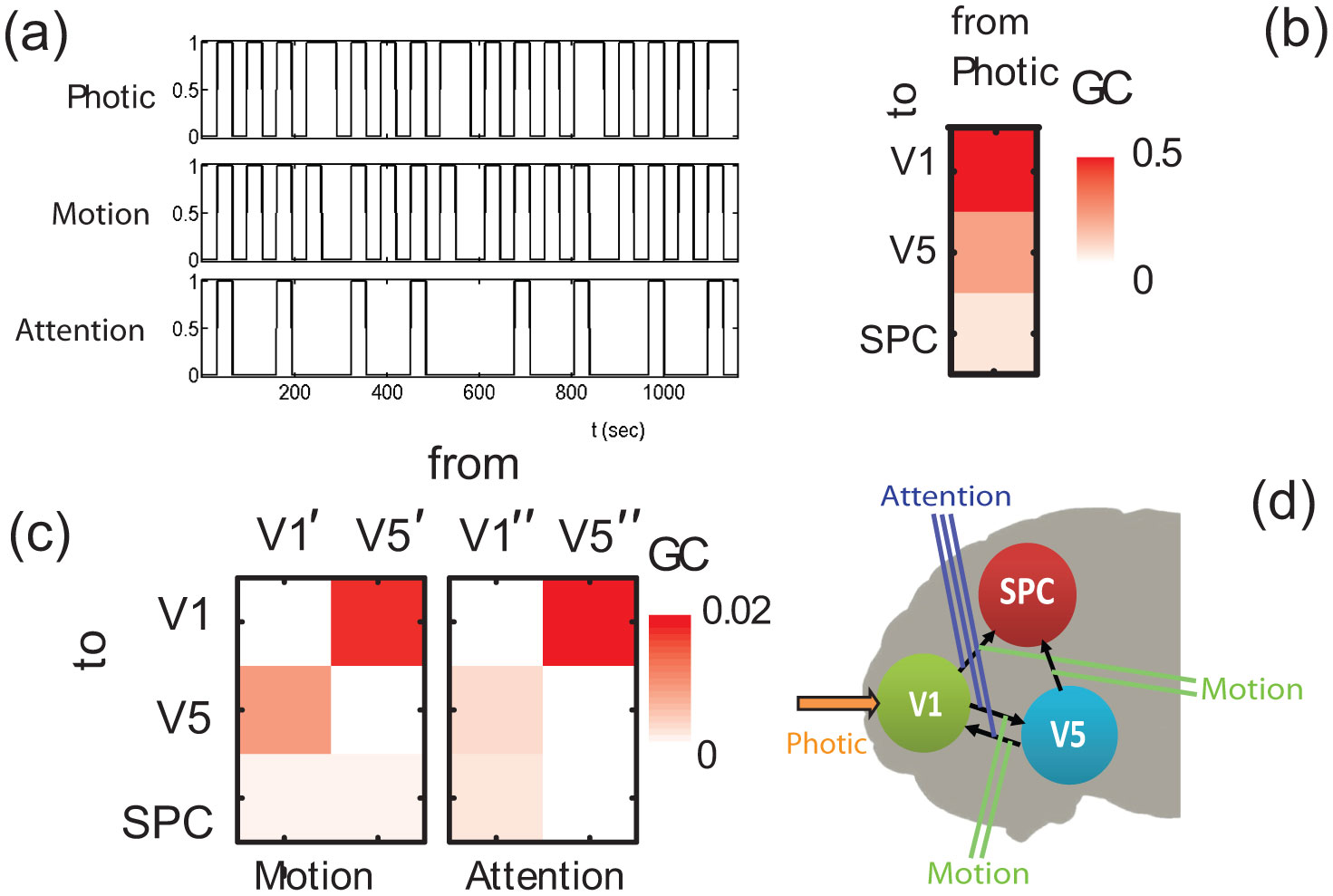
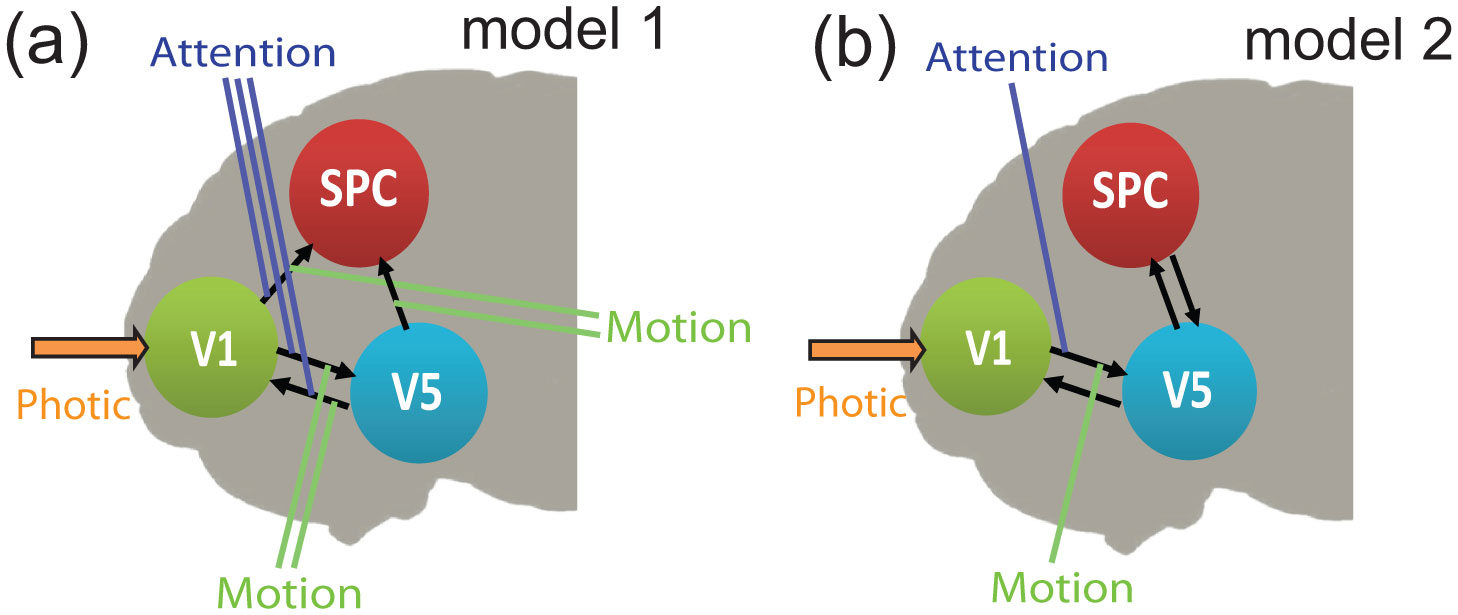
Figures(8) / Tables(2)

Evangelos Almpanis, Constantinos Siettos. Construction of functional brain connectivity networks from fMRI data with driving and modulatory inputs: an extended conditional Granger causality approach[J]. AIMS Neuroscience, 2020, 7(2): 66-88. doi: 10.3934/Neuroscience.2020005







 DownLoad:
DownLoad: