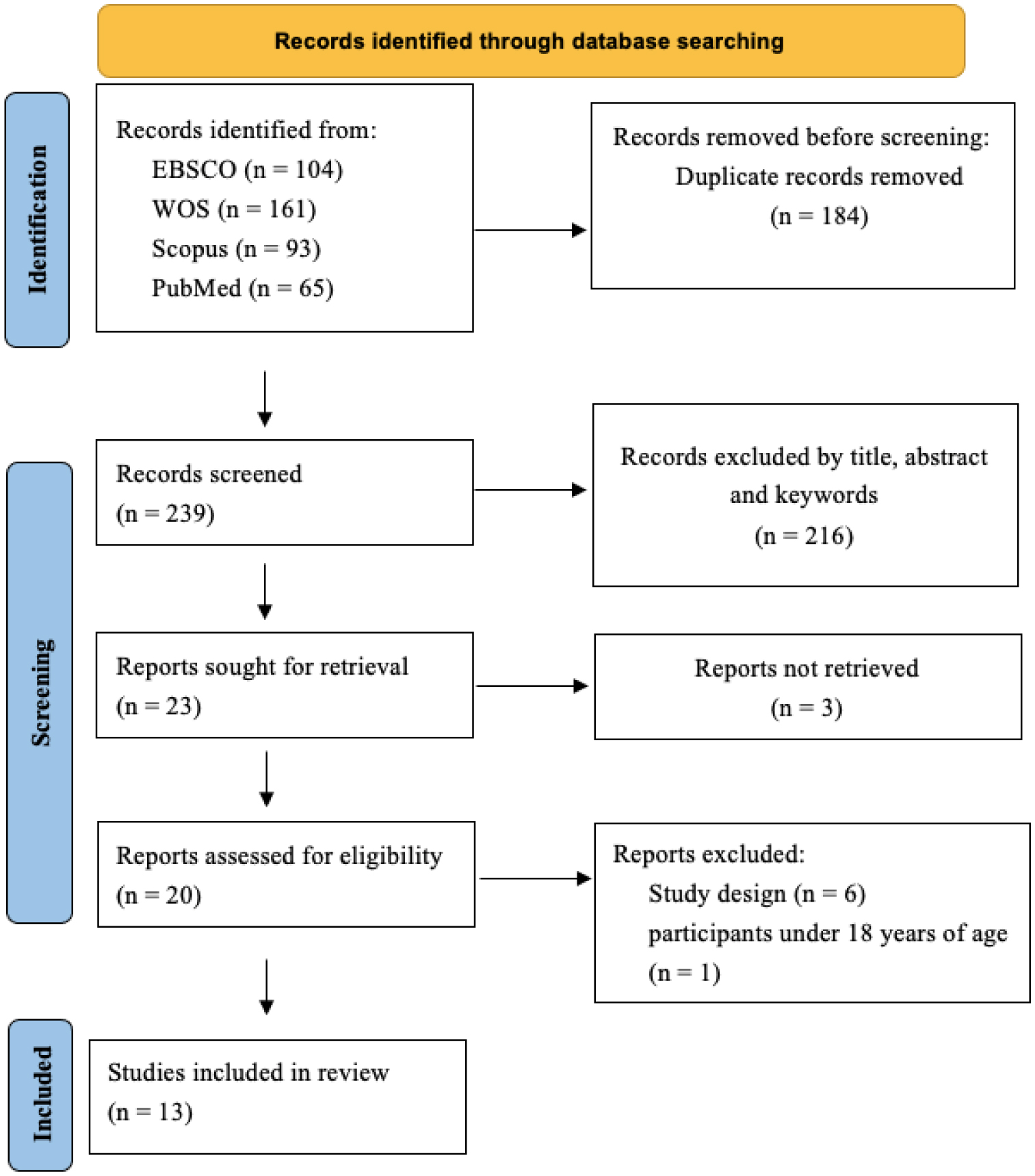
This systematic review aimed to analyze the effects of exercise, physical activity, and sports on physical fitness in adults with Down syndrome (DS). A literature search was conducted across four databases EBSCO, Scopus, Web of Science, and PubMed. The PRISMA guidelines were followed. The PEDro scale and the Cochrane risk of bias tool were used to assess the quality and risk of the studies, respectively. The protocol was registered in PROSPERO (code: CRD42023449627). Of the 423 records initially found, 13 were finally included in the systematic review, in which 349 adults with DS participated. 92% of the articles declared at least one significant difference post-intervention. The available evidence indicates that exercise, physical activity, and sports have a positive effect on some variables of physical fitness, especially strength, balance, body composition, cardiorespiratory fitness, flexibility, and functional capacity. Furthermore, it should be considered as an additional treatment or complementary therapy to improve the functionality and quality of life of adults with DS.
Citation: Felipe Montalva-Valenzuela, Antonio Castillo-Paredes, Claudio Farias-Valenzuela, Oscar Andrades-Ramirez, Yeny Concha-Cisternas, Eduardo Guzmán-Muñoz. Effects of exercise, physical activity, and sports on physical fitness in adults with Down syndrome: A systematic review[J]. AIMS Public Health, 2024, 11(2): 577-600. doi: 10.3934/publichealth.2024029
This systematic review aimed to analyze the effects of exercise, physical activity, and sports on physical fitness in adults with Down syndrome (DS). A literature search was conducted across four databases EBSCO, Scopus, Web of Science, and PubMed. The PRISMA guidelines were followed. The PEDro scale and the Cochrane risk of bias tool were used to assess the quality and risk of the studies, respectively. The protocol was registered in PROSPERO (code: CRD42023449627). Of the 423 records initially found, 13 were finally included in the systematic review, in which 349 adults with DS participated. 92% of the articles declared at least one significant difference post-intervention. The available evidence indicates that exercise, physical activity, and sports have a positive effect on some variables of physical fitness, especially strength, balance, body composition, cardiorespiratory fitness, flexibility, and functional capacity. Furthermore, it should be considered as an additional treatment or complementary therapy to improve the functionality and quality of life of adults with DS.

| [1] |
Franceschi C, Garagnani P, Gensous N, et al. (2019) Accelerated bio-cognitive aging in Down syndrome: State of the art and possible deceleration strategies. Aging Cell 18: e12903. https://doi.org/10.1111/acel.12903 
|
| [2] |
Antonarakis S, Skotko BG, Rafii M, et al. (2020) Down syndrome. Nat Rev Dis Primers 6: 9. https://doi.org/10.1038/s41572-019-0143-7
|
| [3] |
De Graaf G, Buckley F, Skotko BG (2015) Estimates of the live births, natural losses, and elective terminations with Down syndrome in the United States. Am J Med Genet A 167: 756-767. https://doi.org/10.1002/ajmg.a.37001
|
| [4] |
Matson JL, Hess JA, Sipes M, et al. (2010) Developmental profiles from the Battelle developmental inventory: A comparison of toddlers diagnosed with Down Syndrome, global developmental delay and premature birth. Dev Neurorehabil 13: 234-238. https://doi.org/10.3109/17518421003736032
|
| [5] |
Heller T, Hsieh K, Rimmer J (2003) Barriers and supports for exercise participation among adults with Down syndrome. J Gerontol Soc Work 38: 161-178. https://doi.org/10.1300/J083v38n01_0
|
| [6] |
Castro-Piñero J, Carbonell-Baeza A, Martinez-Gomez D, et al. (2014) Follow-up in healthy schoolchildren and in adolescents with Down syndrome: psycho-environmental and genetic determinants of physical activity and its impact on fitness, cardiovascular diseases, inflammatory biomarkers and mental health; the UP&DOWN study. BMC Public Health 14: 1-12. https://doi.org/10.1186/1471-2458-14-400
|
| [7] |
Guzmán-Muñoz EE, Gutiérrez-Navarro LB, Miranda-Díaz SE (2017) Postural control in children, adolescents and adults with Down syndrome. Int Med Rev Down Syndr 21: 12-16. https://doi.org/10.1016/j.sdeng.2016.09.003
|
| [8] |
Ram G, Chinen J (2011) Infections and immunodeficiency in Down syndrome. Clin Exp Immunol 164: 9-16. https://doi.org/10.1111/j.1365-2249.2011.04335.x
|
| [9] | Plaiasu V (2017) Down syndrome–genetics and cardiogenetics. Maedica 12: 208-213. |
| [10] |
Benhaourech S, Drighil A, Hammiri AE (2016) Congenital heart disease and Down syndrome: various aspects of a confirmed association. Cardiovasc J Afr 27: 287-290. https://doi.org/10.5830/CVJA-2016-019
|
| [11] |
Capone GT, Chicoine B, Bulova P, et al. (2018) Co-occurring medical conditions in adults with Down syndrome: A systematic review toward the development of health care guidelines. Am J Med Genet A 176: 116-133. https://doi.org/10.1002/ajmg.a.38512
|
| [12] |
Melo GLR, Neto IVDS, Fonseca EFD, et al. (2022) Resistance training and Down Syndrome: A narrative review on considerations for exercise prescription and safety. Front physiol 13: 948439. https://doi.org/10.3389/fphys.2022.948439
|
| [13] |
Oviedo GR, Carbó-Carreté M, Guerra-Balic M, et al. (2022) Hemodynamic and cardiorespiratory responses to submaximal and maximal exercise in adults with Down syndrome. Front Physiol 13: 905795. https://doi.org/10.3389/fphys.2022.905795
|
| [14] |
Phillips AC, Holland AJ (2011) Assessment of objectively measured physical activity levels in individuals with intellectual disabilities with and without Down's syndrome. PLoS One 6: e28618. https://doi.org/10.1371/journal.pone.0028618
|
| [15] |
Nordstrøm M, Hansen BH, Paus B, et al. (2013) Accelerometer-determined physical activity and walking capacity in persons with Down syndrome, Williams syndrome and Prader–Willi syndrome. Res Dev Disabil 34: 4395-4403. https://doi.org/10.1016/j.ridd.2013.09.021
|
| [16] |
Dairo YM, Collett J, Dawes H, et al. (2016) Physical activity levels in adults with intellectual disabilities: A systematic review. Prev med rep 4: 209-219. https://doi.org/10.1016/j.pmedr.2016.06.008
|
| [17] |
Oreskovic NM, Cottrell C, Torres A, et al. (2020) Physical activity patterns in adults with Down Syndrome. J Appl Res Intellect Disabil 33: 1457-1464. https://doi.org/10.1111/jar.12773
|
| [18] |
Coelho-Junior HJ, Villani ER, Calvani R, et al. (2019) Sarcopenia-related parameters in adults with Down syndrome: a cross-sectional exploratory study. Exp Gerontol 119: 93-99. https://doi.org/10.1016/j.exger.2019.01.028
|
| [19] |
Pitchford EA, Adkins C, Hasson RE, et al. (2018) Association between physical activity and adiposity in adolescents with Down syndrome. Med Sci Sports Exerc 50: 667. https://doi.org/10.1249/MSS.0000000000001502
|
| [20] |
Zhang X, Xie X, Dou Q, et al. (2019) Association of sarcopenic obesity with the risk of all-cause mortality among adults over a broad range of different settings: a updated meta-analysis. BMC Geriatrics 19: 1-14. https://doi.org/10.1186/s12877-019-1195-y
|
| [21] | Corbin CB, Le Masurier GC (2014) Fitness for life, 6 ed. Champaign IL: Human Kinetics. |
| [22] |
Knapik JJ (2015) The importance of physical fitness for injury prevention: part 1. J Spec Oper Med 15: 123-127.
|
| [23] |
Bi C, Zhang F, Gu Y, et al. (2020) Secular trend in the physical fitness of Xinjiang children and adolescents between 1985 and 2014. Int J Environ Res Public Health 17: 2195. https://doi.org/10.3390/ijerph17072195
|
| [24] |
Evaristo S, Moreira C, Lopes L, et al. (2019) Muscular fitness and cardiorespiratory fitness are associated with health-related quality of life: Results from labmed physical activity study. J Exerc Sci Fit 17: 55-61. https://doi.org/10.1016/j.jesf.2019.01.002
|
| [25] |
Solera-Sanchez A, Adelantado-Renau M, Moliner-Urdiales D, et al. (2023) Individual and combined impact of physical fitness on health-related quality of life during adolescence: DADOS Study. Eur J Sport Sci 23: 294-300. https://doi.org/10.1080/17461391.2021.2012596
|
| [26] |
Fardman A, Banschick GD, Rabia R, et al. (2021) Cardiorespiratory fitness is an independent predictor of cardiovascular morbidity and mortality and improves accuracy of prediction models. Can J Cardiol 37: 241-250. https://doi.org/10.1016/j.cjca.2020.05.017
|
| [27] |
García-Hermoso A, Cavero-Redondo I, Ramírez-Vélez R, et al. (2018) Muscular strength as a predictor of all-cause mortality in an apparently healthy population: a systematic review and meta-analysis of data from approximately 2 million men and women. Arch phys med rehab 99: 2100-2113. https://doi.org/10.1016/j.apmr.2018.01.008
|
| [28] |
Chen W, Hammond-Bennett A, Hypnar A, et al. (2018) Health-related physical fitness and physical activity in elementary school students. BMC Public Health 18: 1-12. https://doi.org/10.1186/s12889-018-5107-4
|
| [29] |
Yang J, Christophi CA, Farioli A, et al. (2019) Association between push-up exercise capacity and future cardiovascular events among active adult men. JAMA Net Open 2: e188341. https://doi.org/10.1001/jamanetworkopen.2018.8341
|
| [30] |
Mainous AG, Tanner RJ, Anton SD, et al. (2015) Grip strength as a marker of hypertension and diabetes in healthy weight adults. Am J prev med 49: 850-858. https://doi.org/10.1016/j.amepre.2015.05.025
|
| [31] |
Hao G, Chen H, Ying Y, et al. (2020) The relative handgrip strength and risk of cardiometabolic disorders: a prospective study. Front Physiol 11: 719. https://doi.org/10.3389/fphys.2020.00719
|
| [32] |
Alvarez-Pitti J, Mallén JAC, Trabazo RL, et al. (2020) Ejercicio físico como “medicina” en enfermedades crónicas durante la infancia y la adolescencia. An Pediatr 92: e1-173. https://doi.org/10.1016/j.anpedi.2020.01.010
|
| [33] |
Miko HC, Zillmann N, Ring-Dimitriou S, et al. (2020) Effects of physical activity on health. Gesundheitswesen 82: S184-S195. https://doi.org/10.1055/a-1217-0549
|
| [34] | Legerlotz K (2020) The effects of resistance training on health of children and adolescents with disabilities. Am J Lif Med 14: 382-396. https://doi.org/10.1177/1559827618759640 |
| [35] |
Obrusnikova I, Firkin CJ, Cavalier AR, et al. (2022) Effects of resistance training interventions on muscular strength in adults with intellectual disability: a systematic review and meta-analysis. Dis Rehab 44: 4549-4562. https://doi.org/10.1080/09638288.2021.1910738
|
| [36] |
Obrusnikova I, Firkin CJ, Farquhar WB (2022) A systematic review and meta-analysis of the effects of aerobic exercise interventions on cardiorespiratory fitness in adults with intellectual disability. Dis Health J 15: 101185. https://doi.org/10.1016/j.dhjo.2021.101185
|
| [37] |
Bouzas S, Martínez-Lemos RI, Ayan C (2019) Effects of exercise on the physical fitness level of adults with intellectual disability: A systematic review. Dis Rehab 41: 3118-3140. https://doi.org/10.1080/09638288.2018.1491646
|
| [38] |
Farías-Valenzuela C, Cofré-Bolados C, Ferrari G, et al. (2021) Effects of motor-games-based concurrent training program on body composition indicators of chilean adults with down syndrome. Sustainability 13: 5737. https://doi.org/10.3390/su13105737
|
| [39] |
Maïano C, Hue O, Lepage G, et al. (2019) Do exercise interventions improve balance for children and adolescents with Down syndrome? A systematic review. Phys Ther 99: 507-518. https://doi.org/10.1093/ptj/pzz012
|
| [40] |
Rodríguez-Grande EI, Buitrago-López A, Torres-Narváez MR, et al. (2022) Therapeutic exercise to improve motor function among children with Down Syndrome aged 0 to 3 years: a systematic literature review and meta‑analysis. Sci Rep 12: 13051. https://doi.org/10.1038/s41598-022-16332-x
|
| [41] | Azab AR, Mahmoud WS, Basha MA, et al. (2022) Distinct effects of trampoline-based stretch-shortening cycle exercises on muscle strength and postural control in children with Down syndrome: a randomized controlled study. Eur Rev Med Pharmacol Sci 26: 1952-1962. https://doi.org/10.26355/eurrev_202203_28343 |
| [42] | Barnard M, Swanepoel M, Ellapen TJ, et al. (2019) The health benefits of exercise therapy for patients with Down syndrome: A systematic review. Afr J Dis 8: 1-9. https://doi.org/10.4102/ajod.v8i0.576 |
| [43] | Ballenger BK, Schultz EE, Dale M, et al. (2023) Health outcomes of physical activity interventions in adults with down syndrome: A systematic review. Adap Phys Act Quart 40: 378-402. https://doi.org/10.1123/apaq.2022-0102 |
| [44] | Page MJ, McKenzie JE, Bossuyt PM, et al. (2021) The PRISMA 2020 statement: an updated guideline for reporting systematic reviews. BMJ 29: 372. https://doi.org/10.1136/bmj.n71 |
| [45] |
Valizadeh A, Moassefi M, Nakhostin-Ansari A, et al. (2022) Abstract screening using the automated tool Rayyan: results of effectiveness in three diagnostic test accuracy systematic reviews. BMC Med Resear Methodol 22: 160. https://doi.org/10.1186/s12874-022-01631-8
|
| [46] | Cascaes da Silva F, Valdivia Arancibia BA, da Rosa Iop R, et al. (2013) Escalas y listas de evaluación de la calidad de estudios científicos. Rev Cub Inf Ciencias Salud 24: 295-312. |
| [47] |
De Morton NA (2009) The PEDro scale is a valid measure of the methodological quality of clinical trials: a demographic study. Aust J Physiother 55: 129-133. https://doi.org/10.1016/s0004-9514(09)70043-1
|
| [48] |
Jørgensen L, Paludan-Müller AS, Laursen DR, et al. (2016) Evaluation of the Cochrane tool for assessing risk of bias in randomized clinical trials: overview of published comments and analysis of user practice in Cochrane and non-Cochrane reviews. System Rev 5: 1-13. https://doi.org/10.1186/s13643-016-0259-8
|
| [49] |
Boer PH (2020) The effect of 8 weeks of freestyle swim training on the functional fitness of adults with Down syndrome. J Intelle Disabi Res 64: 770-781. https://doi.org/10.1111/jir.12768
|
| [50] |
Boer PH, De Beer Z (2019) The effect of aquatic exercises on the physical and functional fitness of adults with Down syndrome: A non-randomised controlled trial. J Intelle Disabi Res 63: 1453-1463. https://doi.org/10.1111/jir.12687
|
| [51] |
Boer PH, Moss SJ (2016) Effect of continuous aerobic vs. interval training on selected anthropometrical, physiological and functional parameters of adults with Down syndrome. J Intelle Disabi Res 60: 322-334. https://doi.org/10.1111/jir.12251
|
| [52] |
Cai W, Baek SS (2022) Effects of 24-week basketball programme on body composition and functional fitness on adults with Down syndrome. J Intelle Disabi Res 66: 939-951. https://doi.org/10.1111/jir.12951
|
| [53] |
Cowley PM, Ploutz-Snyder LL, Baynard T, et al. (2011) The effect of progressive resistance training on leg strength, aerobic capacity and functional tasks of daily living in persons with Down syndrome. Disabil Rehabil 33: 2229-2236. https://doi.org/10.3109/09638288.2011.563820
|
| [54] |
Diaz AJ, Rosety I, Ordonez FJ, et al. (2021) Effects of resistance training in muscle mass and markers of muscle damage in adults with down syndrome. Int J Environ Res Public Health 18: 8996. https://doi.org/10.3390/ijerph18178996
|
| [55] |
Perrot A, Maillot P, Le Foulon A, et al. (2021) Effect of exergaming on physical fitness, functional mobility, and cognitive functioning in adults with Down syndrome. Am J Intellect Dev Disabil 126: 34-44. https://doi.org/10.1352/1944-7558-126.1.34
|
| [56] |
Rimmer JH, Heller T, Wang E, et al. (2004) Improvements in physical fitness in adults with Down syndrome. Am J Ment Retard 109: 165-174. https://doi.org/10.1352/0895-8017(2004)109<165:IIPFIA>2.0.CO;2
|
| [57] |
Shields N, Taylor NF, Dodd KJ (2008) Effects of a community-based progressive resistance training program on muscle performance and physical function in adults with Down syndrome: a randomized controlled trial. Arch Phys Med Rehabil 89: 1215-1220. https://doi.org/10.1016/j.apmr.2007.11.056
|
| [58] |
Shin YA, Hong SM, Lee JS, et al. (2021) The effect of resistance and balance training on postural control and physical fitness in adults with down syndrome. Exerc Sci 30: 175-182. https://doi.org/10.15857/ksep.2021.30.2.175
|
| [59] |
Silva V, Campos C, Sá A, et al. (2017) Wii-based exercise program to improve physical fitness, motor proficiency and functional mobility in adults with Down syndrome. J Intelle Disabi Res 61: 755-765. https://doi.org/10.1111/jir.12384
|
| [60] | Tsimaras V, Giagazoglou P, Fotiadou E, et al. (2003) Jog-walk training in cardiorespiratory fitness of adults with Down syndrome. Percept Mot Skills 96: 1239-1251. https://doi.org/10.2466/pms.2003.96.3c.1239 |
| [61] |
Varela AM, Bettencount Sardinha L, Pitetti KH (2001) Effects of an aerobic rowing training regimen in young adults with Down syndrome. Am J Ment Retard 106: 135-144. https://doi.org/10.1352/0895-8017(2001)106<0135:EOAART>2.0.CO;2
|
| [62] |
Martínez-Espinosa RM, Molina Vila MD, Reig Garcia-Galbis M (2020) Evidences from clinical trials in Down syndrome: diet, exercise and body composition. Int J Environ Res Public Health 17: 4294. https://doi.org/10.3390/ijerph17124294
|
| [63] |
Rossato M, Dellagrana RA, da Costa RM, et al. (2018) The accuracy of anthropometric equations to assess body fat in adults with Down syndrome. J Appl Res Intellect Disabil 31: 193-199. https://doi.org/10.1111/jar.12290
|
| [64] |
González-Agüero A, Matute-Llorente Á, Gómez-Cabello A, et al. (2017) Percentage of body fat in adolescents with Down syndrome: Estimation from skinfolds. Disabil Health J 10: 100-104. https://doi.org/10.1016/j.dhjo.2016.05.013
|
| [65] |
Seron BB, Silva RAC, Greguol M (2014) Effects of two programs of exercise on body composition of adolescents with Down syndrome. R Paul Pediatr 32: 92-98. https://doi.org/10.1590/s0103-05822014000100015
|
| [66] |
Holmes CJ, Racette SB (2021) The utility of body composition assessment in nutrition and clinical practice: an overview of current methodology. Nutrients 13: 2493. https://doi.org/10.3390/nu13082493
|
| [67] |
Mikkola TM, Kautiainen H, von Bonsdorff MB, et al. (2020) Body composition and changes in health-related quality of life in older age: a 10-year follow-up of the Helsinki Birth Cohort Study. Qual Life Res 29: 2039-2050. https://doi.org/10.1007/s11136-020-02453-1
|
| [68] |
González-Agüero A, Vicente-Rodríguez G, Gómez-Cabello A, et al. (2011) A combined training intervention programme increases lean mass in youths with Down syndrome. Res Dev Disabil 32: 2383-2388. https://doi.org/10.1016/j.ridd.2011.07.024
|
| [69] |
Strasser B, Burtscher M (2018) Survival of the fittest: VO2max, a key predictor of longevity. Front Biosci 23: 1505-1516. https://doi.org/10.2741/4657
|
| [70] |
Clausen JS, Marott JL, Holtermann A, et al. (2018) Midlife cardiorespiratory fitness and the long-term risk of mortality: 46 years of follow-up. J Am Coll Cardiol 72: 987-995. https://doi.org/10.1016/j.jacc.2018.06.045
|
| [71] | Tranah GJ, Barnes HN, Cawthon PM, et al. (2023) Expression of mitochondrial oxidative stress response genes in muscle is associated with mitochondrial respiration, physical performance, and muscle mass in the Study of Muscle, Mobility and Aging (SOMMA). medRxiv . https://doi.org/10.1101/2023.11.05.23298108 |
| [72] |
Bahiraei S, Ghaderi M, Sharifian E, et al. (2023) Effects of exercise programs on cardiovascular responses in individuals with down syndrome: A systematic review and meta-analysis. Prev Med Rep 36: 102521. https://doi.org/10.1016/j.pmedr.2023.102521
|
| [73] |
Beck VDY, Wee SO, Lefferts EC, et al. (2022) Comprehensive cardiopulmonary profile of individuals with Down syndrome. J Intelle Disabi Res 66: 978-987. https://doi.org/10.1111/jir.12954
|
| [74] |
Soysal P, Hurst C, Demurtas J, et al. (2021) Handgrip strength and health outcomes: Umbrella review of systematic reviews with meta-analyses of observational studies. J Sport Health Sci 10: 290-295. https://doi.org/10.1016/j.jshs.2020.06.009
|
| [75] |
Cannataro R, Cione E, Bonilla DA, et al. (2022) Strength training in elderly: An useful tool against sarcopenia. Front Sports Act Living 4: 950949. https://doi.org/10.3389/fspor.2022.950949
|
| [76] |
Sugimoto D, Bowen SL, Meehan III, et al. (2016) Effects of neuromuscular training on children and young adults with Down syndrome: systematic review and meta-analysis. Res Dev Disabil 55: 197-206. https://doi.org/10.1016/j.ridd.2016.04.003
|
| [77] |
Post EM, Kraemer WJ, Kackley ML, et al. (2022) The effects of resistance training on physical fitness and neuromotor-cognitive functions in adults with down syndrome. Front Rehabil Sci 3: 927629. https://doi.org/10.3389/fresc.2022.927629
|
| [78] |
Gabriel DA, Kamen G, Frost G (2006) Neural adaptations to resistive exercise: mechanisms and recommendations for training practices. Sports Med 36: 133-149. https://doi.org/10.2165/00007256-200636020-00004
|
| [79] |
Kekäläinen T, Luchetti M, Sutin A, et al. (2023) Functional Capacity and Difficulties in Activities of Daily Living From a Cross-National Perspective. J Aging Health 35: 356-369. https://doi.org/10.1177/08982643221128929
|
| [80] |
Terblanche E, Boer PH (2013) The functional fitness capacity of adults with Down syndrome in South Africa. J Intelle Disabi Res 57: 826-836. https://doi.org/10.1111/j.1365-2788.2012.01594.x
|
| [81] |
Jung HK, Chung E, Lee BH (2017) A comparison of the function, activity and participation and quality of life between down syndrome children and typically developing children. J Phys Ther Sci 29: 1377-1380. https://doi.org/10.1589/jpts.29.1377
|
| [82] |
Rodríguez-Grande EI, Vargas-Pinilla OC, Torres-Narvaez MR, et al. (2022) Neuromuscular exercise in children with Down Syndrome: A systematic review. Sci Rep 12: 14988. https://doi.org/10.1038/s41598-022-19086-8
|
| [83] |
Alizadeh S, Daneshjoo A, Zahiri A, et al. (2023) Resistance training induces improvements in range of motion: a systematic review and meta-analysis. Sports Med 53: 707-722. https://doi.org/10.1007/s40279-022-01804-x
|
| [84] |
Guerrero K, Umagat A, Barton M, et al. (2023) The effect of a telehealth exercise intervention on balance in adults with Down syndrome. J Appl Res Intellect Disabil 36: 385-393. https://doi.org/10.1111/jar.13068
|
| [85] |
Šarabon N, Kozinc Ž (2020) Effects of resistance exercise on balance ability: Systematic review and meta-analysis of randomized controlled trials. Life 10: 284. https://doi.org/10.3390/life10110284
|
| [86] |
Oppewal A, Hilgenkamp TI (2019) Physical fitness is predictive for 5-year survival in older adults with intellectual disabilities. J Appl Res Intellect Disabil 32: 958-966. https://doi.org/10.1111/jar.12589
|
| [87] |
Hardee JP, Fetters L (2017) The effect of exercise intervention on daily life activities and social participation in individuals with Down syndrome: A systematic review. Res Dev Disabil 62: 81-103. https://doi.org/10.1016/j.ridd.2017.01.011
|
| [88] |
Alba-Rueda A, Moral-Munoz JA, De Miguel-Rubio, et al. (2022) Exergaming for physical therapy in patients with down syndrome: A systematic review and meta-analysis of randomized-controlled trials. Games Health J 11: 67-78. https://doi.org/10.1089/g4h.2021.0172
|


Figures(3) / Tables(3)

Felipe Montalva-Valenzuela, Antonio Castillo-Paredes, Claudio Farias-Valenzuela, Oscar Andrades-Ramirez, Yeny Concha-Cisternas, Eduardo Guzmán-Muñoz. Effects of exercise, physical activity, and sports on physical fitness in adults with Down syndrome: A systematic review[J]. AIMS Public Health, 2024, 11(2): 577-600. doi: 10.3934/publichealth.2024029







 DownLoad:
DownLoad: