Citation: Amrit Krishna Mitra. Oxytocin and vasopressin: the social networking buttons of the body[J]. AIMS Molecular Science, 2021, 8(1): 32-50. doi: 10.3934/molsci.2021003

| [1] | Acher R, Chauvet J, Chauvet MT (1995) Man and the chimaera. Selective versus neutral oxytocin evolution. Adv Exp Med Biol 395: 615-627. |
| [2] |
Insel TR, Shapiro LE (1992) Oxytocin receptor distribution reflects social organization in monogamous and polygamous voles. Proc Natl Acad Sci USA 89: 5981-5985. doi: 10.1073/pnas.89.13.5981

|
| [3] | Williams JR, Insel TR, Harbaugh CR, et al. (1994) Oxytocin administered centrally facilitates formation of a partner preference in prairie voles (Microtus ochrogaster). J Neuroendocrinol 6: 247-250. |
| [4] | Barberis C, Mounillac B, Durroux T (1998) Structural bases of vaso-pressin/oxytocin receptor function. J Neuroendocrinol 156: 223-229. |
| [5] |
Insel TR (2010) The challenge of translation in social neuroscience: a review of oxytocin, vasopressin, and affiliative behavior. Neuron 65: 768-779. doi: 10.1016/j.neuron.2010.03.005
|
| [6] |
Zink CF, Stein JL, Kempf L, et al. (2010) Vasopressin modulates medial prefrontal cortex-amygdala circuitry during emotion processing in humans. J Neurosci 30: 7017-7022. doi: 10.1523/JNEUROSCI.4899-09.2010
|
| [7] |
Meyer-Lindenberg A (2008) Impact of prosocial neuropeptides on human brain function. Prog Brain Res 170: 463-470. doi: 10.1016/S0079-6123(08)00436-6
|
| [8] |
Goodson JL, Kingsbury MA (2013) What's in a name? Considerations of homologies and nomenclature for vertebrate social behavior networks. Horm Behav 64: 103-112. doi: 10.1016/j.yhbeh.2013.05.006
|
| [9] |
Consiglio AR, Lucion AB (1996) Lesion of hypothalamic paraventricular nucleus and maternal aggressive behavior in female rats. Physiol Behav 59: 591-596. doi: 10.1016/0031-9384(95)02117-5
|
| [10] |
Bosch OJ, Kromer SA, Brunton PJ, et al. (2004) Release of Oxytocin in the hypothalamic paraventricular nucleus, but not central amygdala or lateral septum in lactating residents and virgin intruders during maternal defense. Neuroscience 124: 439-448. doi: 10.1016/j.neuroscience.2003.11.028
|
| [11] |
Giovenardi M, Padoin MJ, Cadore LP, et al. (1997) Hypothalamic paraventricular nucleus, oxytocin, and maternal aggression in rats. Ann N Y Acad Sci 807: 606-609. doi: 10.1111/j.1749-6632.1997.tb51981.x
|
| [12] |
Loup F, Tribollet E, Dubois-Dauphin M, et al. (1991) Localization of high-affinity binding sites for oxytocin and vasopressin in the human brain. An autoradiographic study. Brain Res 555: 220-232. doi: 10.1016/0006-8993(91)90345-V
|
| [13] |
Taylor SE, Saphire-Bernstein S, Seeman TE (2010) Are Plasma Oxytocin in Women and Plasma Vasopressin in Men Biomarkers of Distressed Pair-Bond Relationships? Psychol Sci 21: 3-7. doi: 10.1177/0956797609356507
|
| [14] |
Landgraf R, Neumann ID (2004) Vasopressin and oxytocin release within the brain: a dynamic concept of multiple and variable modes of neuropeptide communication. Front Neuroendocrinol 25: 150-176. doi: 10.1016/j.yfrne.2004.05.001
|
| [15] |
Huber D, Veinante P, Stoop R (2005) Vasopressin and oxytocin excite distinct neuronal populations in the central amygdala. Science 308: 245-248. doi: 10.1126/science.1105636
|
| [16] | Du Vigneaud V (1954-1955) Hormones of the posterior pituitary gland: Oxytocin and vasopressin. Harvey Lect 50: 1-26. |
| [17] |
Bales K, Carter CS (2003) Developmental exposure to oxytocin facilitates partner preferences in male prairie voles (Microtus ochrogaster). Behav Neurosci 117: 854-859. doi: 10.1037/0735-7044.117.4.854
|
| [18] |
Barger N, Hanson KL, Teffer K, et al. (2014) Evidence for evolutionary specialization in human limbic structures. Front Human Neurosci 8: 277. doi: 10.3389/fnhum.2014.00277
|
| [19] |
Ferguson JN, Aldag JM, Insel TR, et al. (2001) Oxytocin in the medial amygdala is essential for social recognition in the mouse. J Neurosci 21: 8278-8285. doi: 10.1523/JNEUROSCI.21-20-08278.2001
|
| [20] |
Kupfermann I (1979) Modulatory actions of neurotransmitters. Annu Rev Neurosci 2: 447-465. doi: 10.1146/annurev.ne.02.030179.002311
|
| [21] |
Leng G, Ludwig M (2008) Neurotransmitters and peptides: whispered secrets and public announcements. J Physiol 586: 5625-5632. doi: 10.1113/jphysiol.2008.159103
|
| [22] |
Wu F, Yu P, Mao L (2018) Analytical and Quantitative in Vivo Monitoring of Brain Neurochemistry by Electrochemical and Imaging Approaches. ACS Omega 3: 13267-13274. doi: 10.1021/acsomega.8b02055
|
| [23] |
Shariatgorji M, Nilsson A, Goodwin RJ, et al. (2014) Direct targeted quantitative molecular imaging of neurotransmitters in brain tissue sections. Neuron 84: 697-707. doi: 10.1016/j.neuron.2014.10.011
|
| [24] |
Finnema SJ, Scheinin M, Shahid M, et al. (2015) Application of cross-species PET imaging to assess neurotransmitter release in brain. Psychopharmacology 232: 4129-4157. doi: 10.1007/s00213-015-3938-6
|
| [25] |
Mirzaei M, Sawan M (2014) Microelectronics-based biosensors dedicated to the detection of neurotransmitters: A review. Sensors 14: 17981-18008. doi: 10.3390/s141017981
|
| [26] |
Evtugyn GA, Hianik T, Nikoleli GP, et al. (2017) Biosensors for Detection of Neurotransmitters and Neurodegenerative Related Diseases. Front Clin Drug Res: CNS Neurol Disord 5: 184-210. doi: 10.2174/9781681085852117050007
|
| [27] |
Bradford H (1995) Glutamate, GABA and epilepsy. Prog Neurobiol 47: 477-511. doi: 10.1016/0301-0082(95)00030-5
|
| [28] |
Chapman AG (2000) Glutamate and epilepsy. J Nutr 130: 1043S-1045S. doi: 10.1093/jn/130.4.1043S
|
| [29] |
Cavallero A, Marte A, Fedele E (2009) L-Aspartate as an amino acid neurotransmitter: Mechanisms of the depolarization-induced release from cerebrocortical synaptosomes. J Neurochem 110: 924-934. doi: 10.1111/j.1471-4159.2009.06187.x
|
| [30] |
Baughman RW, Gilbert CD (1981) Aspartate and glutamate as possible neurotransmitters in the visual cortex. J Neurosci 1: 427-439. doi: 10.1523/JNEUROSCI.01-04-00427.1981
|
| [31] |
Balthazart J, Ball GF (2006) Is brain estradiol a hormone or a neurotransmitter? Trends Neurosci 29: 241-249. doi: 10.1016/j.tins.2006.03.004
|
| [32] |
Brosens JJ, Tullet J, Varshochi R, et al. (2004) Steroid receptor action. Best Pract Res Clin Obstet Gynaecol 18: 265-283. doi: 10.1016/j.bpobgyn.2004.01.006
|
| [33] |
Feldman R (2012) Oxytocin and social affiliation in humans. Horm Behav 61: 380-391. doi: 10.1016/j.yhbeh.2012.01.008
|
| [34] |
Legros JJ (2001) Inhibitory effect of oxytocin on corticotrope function in humans: are vasopressin and oxytocin ying-yang neurohormones? Psychoneuroendocrinology 26: 649-655. doi: 10.1016/S0306-4530(01)00018-X
|
| [35] |
Bailey DJ, Ma C, Soma KK, et al. (2013) Inhibition of hippocampal aromatization impairs spatial memory performance in a male songbird. Endocrinology 154: 4707-4714. doi: 10.1210/en.2013-1684
|
| [36] |
Dale HH (1906) On some physiological actions of ergot. J Physiol 34: 163-206. doi: 10.1113/jphysiol.1906.sp001148
|
| [37] |
Lee AG, Cool DR, Grunwald WC, et al. (2011) A novel form of oxytocin in New World monkeys. Biol Lett 7: 584-587. doi: 10.1098/rsbl.2011.0107
|
| [38] | Mitra AK (2020) Antioxidants: A Masterpiece of Mother Nature to Prevent Illness. J Chem Rev 2: 243-256. |
| [39] |
du Vigneaud V, Ressler C, Swan JM, et al. (1954) The Synthesis of Oxytocin. J Am Chem Soc 76: 3115-3121. doi: 10.1021/ja01641a004
|
| [40] |
Lim MM, Young LJ (2004) Vasopressin-dependent neural circuits underlying pair bond formation in the monogamous prairie vole. Neuroscience 125: 35-45. doi: 10.1016/j.neuroscience.2003.12.008
|
| [41] |
Salata RA, Jarrett DB, Verbalis JG, et al. (1988) Vasopressin stimulation of adrenocorticotropin hormone (ACTH) in humans. In vivo bioassay of corticotropin-releasing factor (CRF) which provides evidence for CRF mediation of the diurnal rhythm of ACTH. J Clin Invest 81: 766-774. doi: 10.1172/JCI113382
|
| [42] |
Chauvet MT, Hurpet D, Chauvet J, et al. (1980) Phenypressin (Phe2-Arg8-vasopressin), a new neurohypophysial peptide found in marsupials. Nature 287: 640-642. doi: 10.1038/287640a0
|
| [43] |
Iovino M, Messana T, De Pergola G, et al. (2018) The Role of Neurohypophyseal Hormones Vasopressin and Oxytocin in Neuropsychiatric Disorders. Endocr Metab Immune Disord Drug Targets 18: 341-347. doi: 10.2174/1871530318666180220104900
|
| [44] |
Caldwell HK, Lee HJ, Macbeth AH, et al. (2008) Vasopressin: behavioral roles of an “original” neuropeptide. Prog Neurobiol 84: 1-24. doi: 10.1016/j.pneurobio.2007.10.007
|
| [45] |
Macdonald K, Macdonald TM (2010) The peptide that binds: a systematic review of oxytocin and its prosocial effects in humans. Harv Rev Psychiatry 18: 1-21. doi: 10.3109/10673220903523615
|
| [46] |
Burbach JPH, de Hoop MJ, Schmale H, et al. (1984) Differential responses to osmotic stress of vasopressin-neurophysin mRNA in hypo-thalamic nuclei. Neuroendocrinology 39: 582-584. doi: 10.1159/000124040
|
| [47] |
Winslow J, Insel T (2002) The social deficits of the oxytocin knockout mouse. Neuropeptides 36: 221-229. doi: 10.1054/npep.2002.0909
|
| [48] |
Bales KL, Kim AL, Lewis-Reese AD, et al. (2004) Both oxytocin and vasopressin may influence alloparental behavior in male prairie voles. Horm Behav 45: 354-361. doi: 10.1016/j.yhbeh.2004.01.004
|
| [49] |
Saito A, Nakamura K (2011) Oxytocin changes primate paternal tolerance to offspring in food transfer. J Comp Physiol A 197: 329-337. doi: 10.1007/s00359-010-0617-2
|
| [50] |
Bielsky IF, Young LJ (2004) Oxytocin, vasopressin, and social recognition in mammals. Peptides 25: 1565-1574. doi: 10.1016/j.peptides.2004.05.019
|
| [51] |
Odendaal JS, Meintjes RA (2003) Neurophysiological correlates of affiliative behaviour between humans and dogs. Vet J 165: 296-301. doi: 10.1016/S1090-0233(02)00237-X
|
| [52] |
Bick J, Dozier M (2010) Mothers' and Children's Concentrations of Oxytocin Following Close, Physical Interactions with Biological and Non-biological Children. Dev Psychobiol 52: 100-107. doi: 10.1002/dev.20411
|
| [53] |
Sheng F, Liu Y, Zhou B, et al. (2013) Oxytocin modulates the racial bias in neural responses to others' suffering. Biol Psychol 92: 380-386. doi: 10.1016/j.biopsycho.2012.11.018
|
| [54] |
Lane A, Luminet O, Rimé B, et al. (2013) Oxytocin increases willingness to socially share one's emotions. Int J Psychol 48: 676-681. doi: 10.1080/00207594.2012.677540
|
| [55] |
Shamay-Tsoory SG, Fischer M, Dvash J, et al. (2009) Intranasal administration of oxytocin increases envy and schadenfreude (gloating). Biol Psychiatry 66: 864-870. doi: 10.1016/j.biopsych.2009.06.009
|
| [56] |
Tabak BA, McCullough ME, Carver CS, et al. (2014) Variation in oxytocin receptor gene (OXTR) polymorphisms is associated with emotional and behavioral reactions to betrayal. Soc Cogn Affect Neurosci 9: 810-816. doi: 10.1093/scan/nst042
|
| [57] |
Viviani D, Charlet A, van den Burg E, et al. (2011) Oxytocin selectively gates fear responses through distinct outputs from the central amygdala. Science 333: 104-107. doi: 10.1126/science.1201043
|
| [58] |
Okabe S, Kitano K, Nagasawa M, et al. (2013) Testosterone inhibits facilitating effects of parenting experience on parental behavior and the oxytocin neural system in mice. Physiol Behav 118: 159-164. doi: 10.1016/j.physbeh.2013.05.017
|
| [59] |
Blaicher W, Gruber D, Bieglmayer C, et al. (1999) The role of oxytocin in relation to female sexual arousal. Gynecol Obstet Invest 47: 125-126. doi: 10.1159/000010075
|
| [60] |
Anderson-Hunt M, Dennerstein L (1995) Oxytocin and female sexuality. Gynecol Obstet Invest 40: 217-221. doi: 10.1159/000292340
|
| [61] |
Murphy MR, Seckl JR, Burton S, et al. (1987) Changes in oxytocin and vasopressin secretion during sexual activity in men. J Clin Endocrinol Metab 65: 738-741. doi: 10.1210/jcem-65-4-738
|
| [62] |
Krüger TH, Haake P, Chereath D, et al. (2003) Specificity of the neuroendocrine response to orgasm during sexual arousal in men. J Endocrinol 177: 57-64. doi: 10.1677/joe.0.1770057
|
| [63] |
Haas M, Glick SM (1978) Radioimmunoassayable plasma vasopressin associated with surgery. Arch Surg 113: 597-600. doi: 10.1001/archsurg.1978.01370170059011
|
| [64] |
Nussey SS, Page SR, Ang VT, et al. (1988) The response of plasma oxytocin to surgical stress. Clin Endocrinol 28: 277-282. doi: 10.1111/j.1365-2265.1988.tb01213.x
|
| [65] |
Lolait SJ, Stewart LQ, Jessop DS, et al. (2007) The hypothalamic-pituitary-adrenal axis response to stress in mice lacking functional vasopressin V1b receptors. Endocrinology 148: 849-856. doi: 10.1210/en.2006-1309
|
| [66] |
Acher R, Chauvet J (1995) The neurohypophysial endocrine regulatory cascade: precursors, mediators, receptors, and effectors. Front Neuroendocrinol 16: 237-289. doi: 10.1006/frne.1995.1009
|
| [67] | Robertson GL (1975) The regulation of vasopressin functions in health and disease. Recent Prog Horm 33: 333-385. |
| [68] |
Weingartner H, Gold P, Ballenger J, et al. (1981) Effects of vasopressin on human memory functions. Science 211: 601-603. doi: 10.1126/science.7455701
|
| [69] |
Albeck D, Smock T (1988) A mechanism for vasopressin action in the hippocampus. Brain Res 463: 394-397. doi: 10.1016/0006-8993(88)90417-9
|
| [70] |
Yehuda S (1987) Effects of alpha-MSH, TRH and AVP on learning and memory, pain threshold, and motor activity: preliminary results. Int J Neurosci 32: 703-709. doi: 10.3109/00207458709043325
|
| [71] | Mavani GP, DeVita MV, Michelis MF (2015) A Review of the Nonpressor and Nonantidiuretic Actions of the Hormone Vasopressin. Front Med 2: 1-11. |
| [72] |
Hendaus MA, Jomha FA, Alhammadi AH (2019) Vasopressin in the Amelioration of Social Functioning in Autism Spectrum Disorder. J Clin Med 8: 1061. doi: 10.3390/jcm8071061
|
| [73] |
Hall SS, Lightbody AA, Reiss AL (2008) Compulsive, self-injurious, and autistic behavior in children and adolescents with fragile X syndrome. Am J Ment Retard 113: 44-53. doi: 10.1352/0895-8017(2008)113[44:CSAABI]2.0.CO;2
|
| [74] |
Ji N, Findling RL (2015) An update on pharmacotherapy for autism spectrum disorder in children and adolescents. Curr Opin Psychiatry 28: 91-101. doi: 10.1097/YCO.0000000000000132
|
| [75] |
Manning M, Stoev S, Chini B, et al. (2008) Peptide and non-peptide agonists and antagonists for the vasopressin and oxytocin V1a, V1b, V2 and OT receptors: research tools and potential therapeutic agents. Prog Brain Res 170: 473-512. doi: 10.1016/S0079-6123(08)00437-8
|
| [76] |
Porges SW (1998) Love: an emergent property of the mammalian autonomic nervous system. Psychoneuroendocrinology 23: 837-861. doi: 10.1016/S0306-4530(98)00057-2
|
| [77] |
Weisman O, Zagoory-Sharon O, Feldman R (2012) Intranasal oxytocin administration is reflected in human saliva. Psychoneuroendocrinology 37: 1582-1586. doi: 10.1016/j.psyneuen.2012.02.014
|
| [78] | Huffmeijer R, Alink LR, Tops M, et al. (2012) Salivary levels of oxytocin remain elevated for more than two hours after intranasal oxytocin administration. Neuro Endocrinol Lett 33: 21-25. |
| [79] |
Guastella AJ, Mitchell PB, Mathews F (2008) Oxytocin enhances the encoding of positive social memories in humans. Biol Psychiatry 64: 256-258. doi: 10.1016/j.biopsych.2008.02.008
|
| [80] |
Torloni MR, Gomes Freitas C, Kartoglu UH, et al. (2016) Quality of oxytocin available in low- and middle-income countries: a systematic review of the literature. BJOG 123: 2076-2086. doi: 10.1111/1471-0528.13998
|
| [81] | Mehta AC (1986) Buccal and oral drugs: induction of labor. Acta Chirurgica Hungarica 27: 157-163. |
| [82] | Baum S, Nusbaum M, Tumen HJ (1970) The control of gastrointestinal hemorrhage by selective mesenteric infusion of pitressin. Gastroenterology 58: 926. |
| [83] |
Lauzier F, Lévy B, Lamarre P, et al. (2006) Vasopressin or norepinephrine in early hyperdynamic septic shock: a randomized clinical trial. primary. Intensive Care Med 32: 1782-1789. doi: 10.1007/s00134-006-0378-0
|
| [84] |
Aung K, Htay T (2005) Vasopressin for cardiac arrest: a systematic review and meta-analysis. Arch Intern Med 165: 17-24. doi: 10.1001/archinte.165.1.17
|
| [85] |
Dünser MW, Mayr AJ, Ulmer H, et al. (2001) The effects of vasopressin on systemic hemodynamics in catecholamine-resistant septic and postcardiotomy shock: a retrospective analysis. Anesth Analg 93: 7-13. doi: 10.1097/00000539-200107000-00003
|
| [86] |
Savaskan E, Ehrhardt R, Schulz A, et al. (2008) Post-learning intranasal oxytocin modulates human memory for facial identity. Psychoneuroendocrinology 33: 368-374. doi: 10.1016/j.psyneuen.2007.12.004
|
| [87] |
Lischke A, Berger C, Prehn K, et al. (2012) Intranasal oxytocin enhances emotion recognition from dynamic facial expressions and leaves eye-gaze unaffected. Psychoneuroendocrinology 37: 475-481. doi: 10.1016/j.psyneuen.2011.07.015
|
| [88] |
Dal Monte O, Noble PL, Costa VD, et al. (2014) Oxytocin enhances attention to the eye region in rhesus monkeys. Front Neurosci 8: 41. doi: 10.3389/fnins.2014.00041
|
| [89] |
Chang SW, Barter JW, Ebitz RB, et al. (2012) Inhaled oxytocin amplifies both vicarious reinforcement and self reinforcement in rhesus macaques (Macaca mulatta). Proc Natl Acad Sci 109: 959-964. doi: 10.1073/pnas.1114621109
|
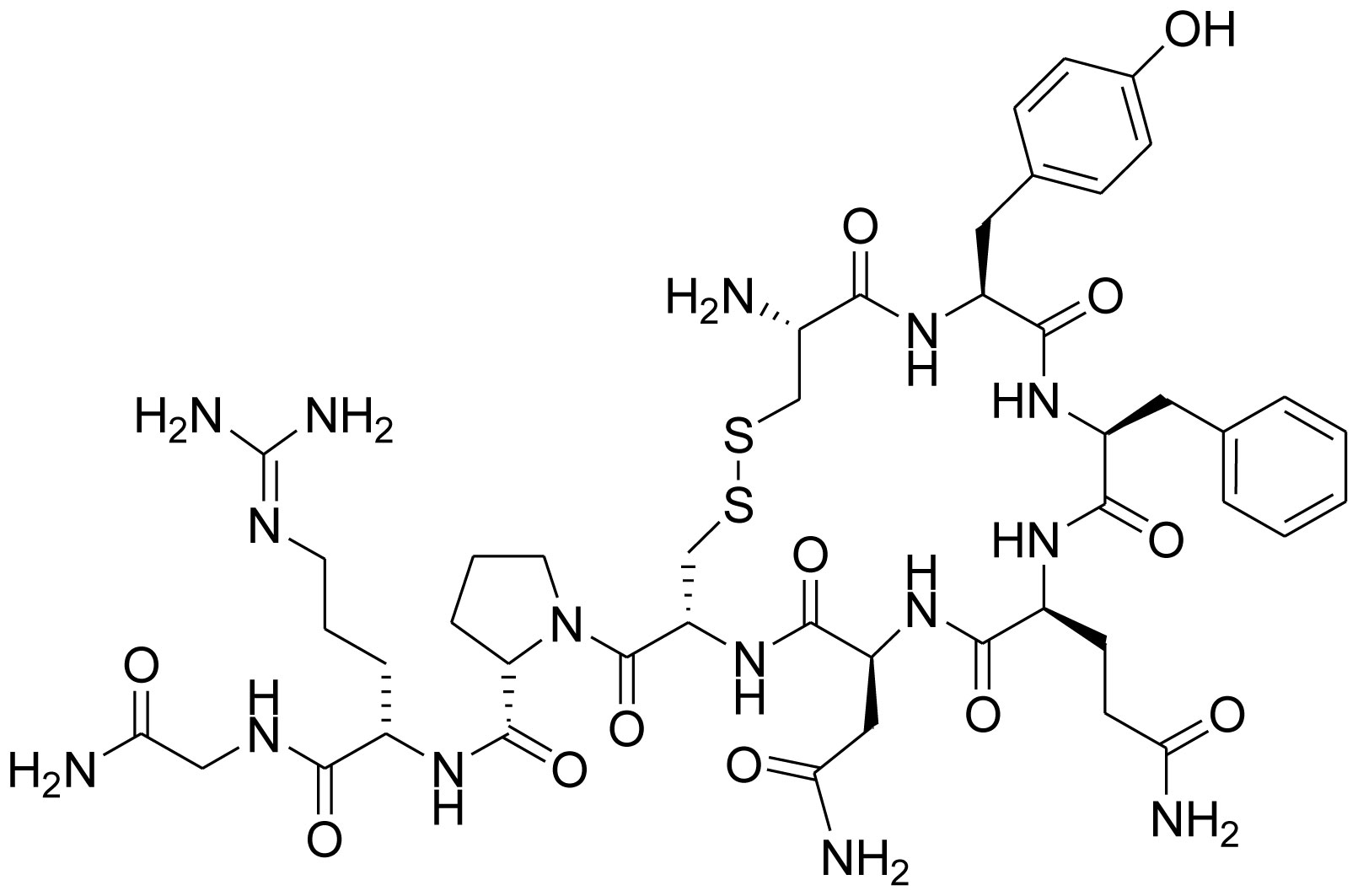
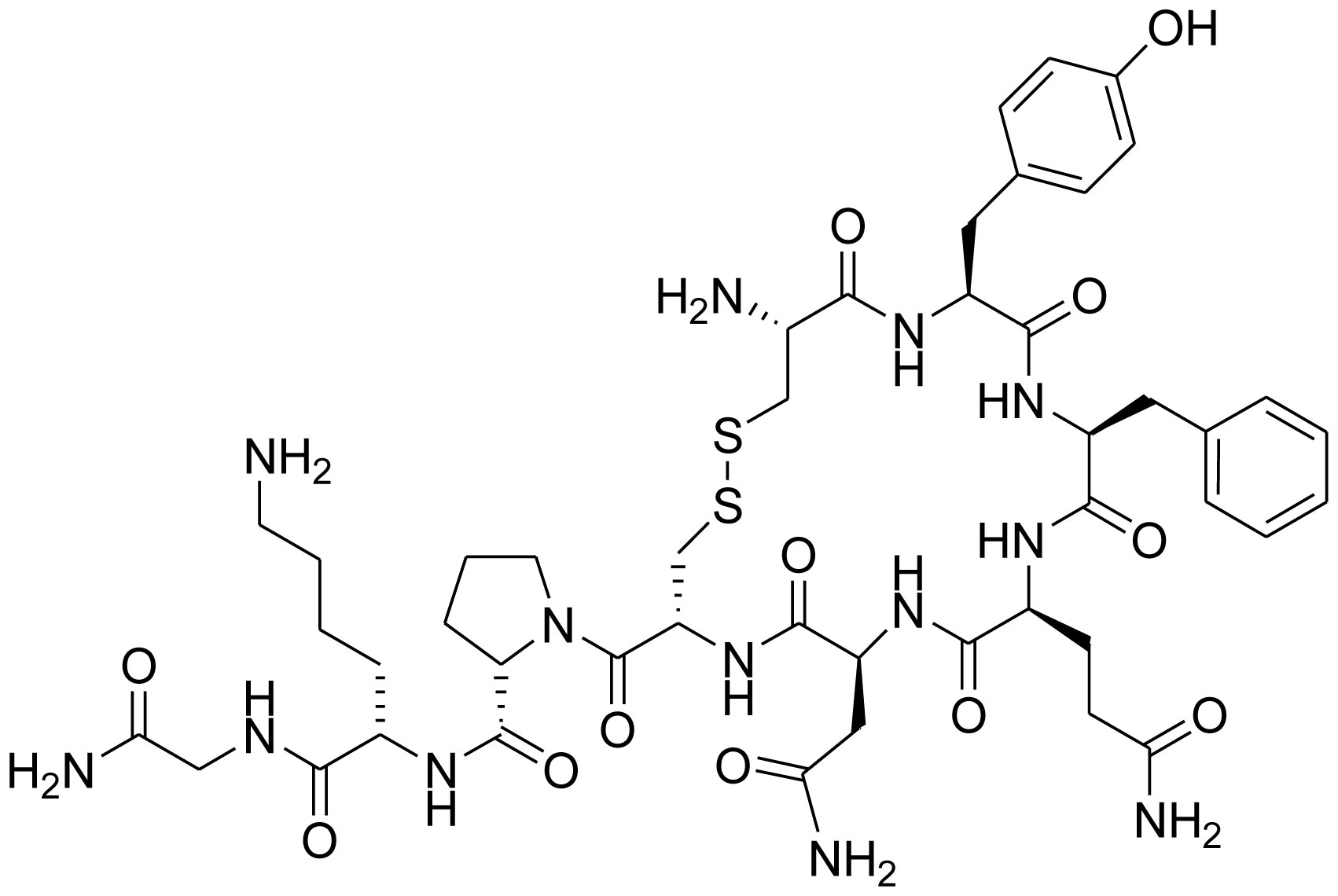
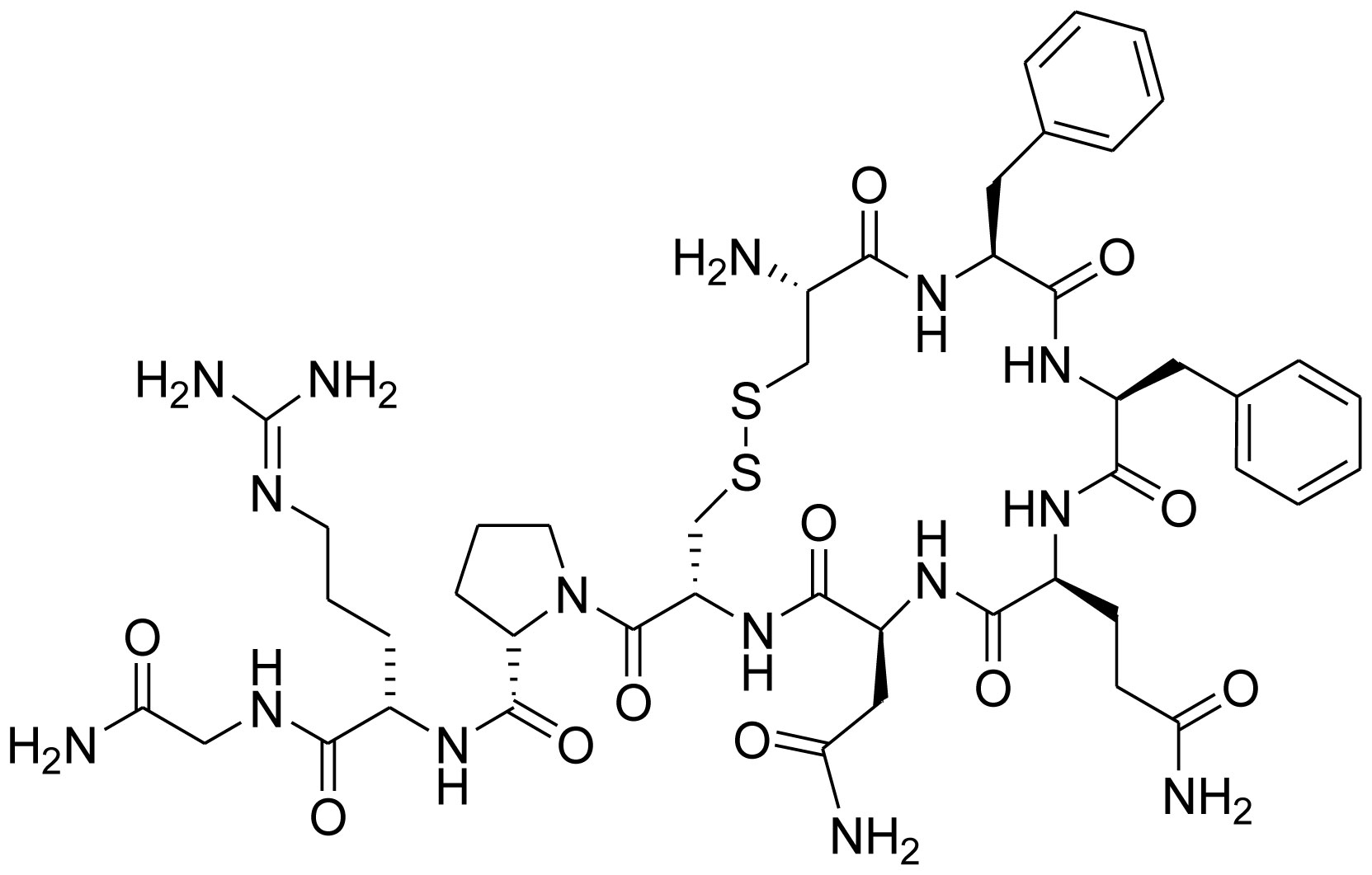
Figures(7) / Tables(1)

Amrit Krishna Mitra. Oxytocin and vasopressin: the social networking buttons of the body[J]. AIMS Molecular Science, 2021, 8(1): 32-50. doi: 10.3934/molsci.2021003







 DownLoad:
DownLoad: