In ecology, the impact of predators goes beyond killing prey, the mere presence of predators reduces the ability of prey to reproduce. In this study, we extend the predator-prey model with fear effect by introducing the state-dependent control with a nonlinear action threshold depending on the combination of the density of prey and its changing rate. We initially defined the Poincaré map of the proposed model and studied its fundamental properties. Utilizing the properties of the Poincaré map, periodic solution of the model is further investigated, including the existence and stability of the order-1 periodic solution and the existence of the order-k ($ k \ge 2 $) periodic solutions. In addition, the influence of the fear effect on the system's dynamics is explored through numerical simulations. The action threshold used in this paper is more consistent with the actual growth of the population than in earlier linear threshold studies, and the results show that the control objectives are better achieved using the action threshold strategy. The analytical approach used in this study provided several novel methods for analyzing the complex dynamics that rely on state-dependent impulsive.
Citation: Yazhi Wu, Guangyao Tang, Changcheng Xiang. Dynamic analysis of a predator-prey state-dependent impulsive model with fear effect in which action threshold depending on the prey density and its changing rate[J]. Mathematical Biosciences and Engineering, 2022, 19(12): 13152-13171. doi: 10.3934/mbe.2022615
In ecology, the impact of predators goes beyond killing prey, the mere presence of predators reduces the ability of prey to reproduce. In this study, we extend the predator-prey model with fear effect by introducing the state-dependent control with a nonlinear action threshold depending on the combination of the density of prey and its changing rate. We initially defined the Poincaré map of the proposed model and studied its fundamental properties. Utilizing the properties of the Poincaré map, periodic solution of the model is further investigated, including the existence and stability of the order-1 periodic solution and the existence of the order-k ($ k \ge 2 $) periodic solutions. In addition, the influence of the fear effect on the system's dynamics is explored through numerical simulations. The action threshold used in this paper is more consistent with the actual growth of the population than in earlier linear threshold studies, and the results show that the control objectives are better achieved using the action threshold strategy. The analytical approach used in this study provided several novel methods for analyzing the complex dynamics that rely on state-dependent impulsive.

| [1] |
A. Hastings, T. Powell, Chaos in a three-species food chain, Ecology, 72 (1991), 896–903. http://dx.doi.org/https://doi.org/10.2307/1940591 doi: 10.2307/1940591

|
| [2] |
M. F. Danca, J. Chattopadhyay, Chaos control of hastings-powell model by combining chaotic motions, Chaos Interdiscipl. J. Nonlinear Sci., 26 (2016), 043106. http://dx.doi.org/https://doi.org/10.1063/1.4946811 doi: 10.1063/1.4946811
|
| [3] |
G. Maciorowski, U. Jankowiak, T. H. Sparks, M. Polakowski, P. Tryjanowski, Biodiversity hotspots at a small scale: The importance of eagles' nests to many other animals, Bullet. Ecolog. Soc. Am., 102 (2021), e03220. http://dx.doi.org/10.1002/ecy.3220 doi: 10.1002/ecy.3220
|
| [4] | J. Robert, Predation, Taylor, Springer Netherlands, 1984. http://dx.doi.org/http://dx.doi.org/10.1179/136217103225005633 |
| [5] |
S. L. Lima, Nonlethal effects in the ecology of predator-prey interactions, Bioscience, 48 (1998), 25–34. http://dx.doi.org/10.2307/1313225 doi: 10.2307/1313225
|
| [6] | W. B. Cannon, Bodily changes in pain, hunger, fear and rage, Bodily changes in pain, hunger, fear, and rage, 1915. http://dx.doi.org/10.1037/10013-000 |
| [7] |
X. Wang, L. Zanette, X. Zou, Modelling the fear effect in predator-prey interactions, J. Math. Biol., 73 (2016), 1–26. http://dx.doi.org/10.1007/s00285-016-0989-1 doi: 10.1007/s00285-016-0989-1
|
| [8] |
X. Wang, X. Zou, Modeling the fear effect in predatorey interactions with adaptive avoidance of predators, Bullet. Math. Biol., 79 (2017), 1–35. http://dx.doi.org/10.1007/s11538-017-0287-0 doi: 10.1007/s11538-017-0287-0
|
| [9] |
L. Y. Zanette, A. F. White, M. C. Allen, M. Clinchy, Perceived predation risk reduces the number of offspring songbirds produce per year, Science, 334 (2011), 1398–1401. http://dx.doi.org/https://doi.org/10.1126/science.1210908 doi: 10.1126/science.1210908
|
| [10] |
A. Das, G. P. Samanta, Modelling the fear effect on a stochastic prey-predator system with additional food for predator, J. Phys. A Math. Theoret., 51 (2018), 465–601. http://dx.doi.org/10.1088/1751-8121/aae4c6 doi: 10.1088/1751-8121/aae4c6
|
| [11] | D. Sahoo, G. P. Samanta, Impact of fear effect in a two prey-one predator system with switching behaviour in predation, Differ. Equat. Dynam. Syst., (2021), 1–23. http://dx.doi.org/https://doi.org/10.1007/s12591-021-00575-7 |
| [12] |
B. K. Das, D. Sahoo, G. P. Samanta, Impact of fear in a delay-induced predator-prey system with intraspecific competition within predator species, Math. Comput. Simul. (MATCOM), 191 (2022), 134–156. http://dx.doi.org/10.1016/j.matcom.2021.08.005 doi: 10.1016/j.matcom.2021.08.005
|
| [13] |
K. Sarkar, S. Khajanchi, Impact of fear effect on the growth of prey in a predator-prey interaction model, Ecolog. Complex., 42 (2020), 100826. http://dx.doi.org/10.1016/j.ecocom.2020.100826 doi: 10.1016/j.ecocom.2020.100826
|
| [14] |
V. Kumar, N. Kumari, Stability and bifurcation analysis of hassell-varley prey-predator system with fear effect, Int. J. Appl. Comput. Math., 6 (2020), 1–20. http://dx.doi.org/10.1007/s40819-020-00899-y doi: 10.1007/s40819-020-00899-y
|
| [15] | S. A. Rahi, S. Kurnaz, R. K. Naji, The impact of fear on a stage structure predator system with anti-predator behavior, Appl. Nanosci., (2020), 1–16. http://dx.doi.org/10.1007/s13204-021-02160-4 |
| [16] |
N. Pati, S. Garai, M. Hossain, G. C. Layek, N. Pal, Fear induced multistability in a predator-prey model, Int. J. Bifurc. Chaos, 31 (2021), 2150150. https://doi.org/10.1142/s0218127421501509 doi: 10.1142/s0218127421501509
|
| [17] |
I. U. Khan, S. Tang, B. Tang, The state-dependent impulsive model with action threshold depending on the pest density and its changing rate, Complexity, 2019 (2019), 6509867. http://dx.doi.org/10.1155/2019/6509867 doi: 10.1155/2019/6509867
|
| [18] | W. Zhao, J. Li, X. Meng, Dynamical analysis of SIR epidemic model with nonlinear pulse vaccination and lifelong immunity, Discrete Dynam. Nat. Soc., 2015. http://dx.doi.org/10.1155/2015/848623 |
| [19] |
S. Tang, B. Tang, A. Wang, Y. Xiao, Holling II predator-prey impulsive semi-dynamic model with complex Poincaré map, Nonlinear Dynam., 81 (2015), 1575–1596. http://dx.doi.org/10.1007/s11071-015-2092-3 doi: 10.1007/s11071-015-2092-3
|
| [20] |
X. Meng, L. Zhang, Evolutionary dynamics in a lotka-volterra competition model with impulsive periodic disturbance, Math. Methods Appl. Sci., 39 (2016), 177–188. http://dx.doi.org/10.1002/mma.3467 doi: 10.1002/mma.3467
|
| [21] |
X. Yu, S. Yuan, T. Zhang, Asymptotic properties of stochastic nutrient-plankton food chain models with nutrient recycling, Nonlinear Anal. Hybrid Syst., 34 (2019), 209–225. http://dx.doi.org/10.1016/j.nahs.2019.06.005 doi: 10.1016/j.nahs.2019.06.005
|
| [22] |
D. Fang, Y. Pei, Y. Lv, L. Chen, Periodicity induced by state feedback controls and driven by disparate dynamics of a herbivore–plankton model with cannibalism, Nonlinear Dynam., 90 (2017), 2657–2672. http://dx.doi.org/10.1007/s11071-017-3829-y doi: 10.1007/s11071-017-3829-y
|
| [23] | D. Li, Y. Liu, H. Cheng, Dynamic complexity of a phytoplankton-fish model with the impulsive feedback control by means of poincarmap, Complexity, 2020. http://dx.doi.org/10.1155/2020/8974763 |
| [24] |
J. Yang, S. Tang, Holling type II predator-prey model with nonlinear pulse as state-dependent feedback control, Elsevier Sci. Publishers B. V., 291 (2016), 225–241. http://dx.doi.org/10.1016/j.cam.2015.01.017 doi: 10.1016/j.cam.2015.01.017
|
| [25] |
Z. Shi, H. Cheng, Y. Liu, Y. Wang, Optimization of an integrated feedback control for a pest management predator-prey model, Math. Biosci. Eng., 16 (2019), 7963–7981. http://dx.doi.org/10.3934/mbe.2019401 doi: 10.3934/mbe.2019401
|
| [26] |
I. U. Khan, S. Tang, The impulsive model with pest density and its change rate dependent feedback control, Discrete Dynam. Nat. Soc., 2020 (2020), 1–20. http://dx.doi.org/10.1155/2020/4561241 doi: 10.1155/2020/4561241
|
| [27] |
T. Li, W. Zhao, Periodic solution of a neutral delay leslie predator-prey model and the effect of random perturbation on the smith growth model, Complexity, 2020 (2020), 1–15. http://dx.doi.org/10.1155/2020/8428269 doi: 10.1155/2020/8428269
|
| [28] |
Y. Li, Y. Li, Y. Liu, H. Cheng, Stability analysis and control optimization of a prey-predator model with linear feedback control, Discrete Dynam. Nat. Soc., 2018 (2018), 1–12. http://dx.doi.org/10.1155/2018/4945728 doi: 10.1155/2018/4945728
|
| [29] |
Z. Shi, H. Cheng, Y. Liu, Y. Li, A cydia pomonella integrated management predator-prey model with smith growth and linear feedback control, IEEE Access, 7 (2019), 126066–126076. http://dx.doi.org/10.1109/ACCESS.2019.2938772 doi: 10.1109/ACCESS.2019.2938772
|
| [30] |
H. Qi, X. Leng, X. Meng, T. Zhang, Periodic solution and ergodic stationary distribution of seis dynamical systems with active and latent patients, Qual. Theory Dynam. Syst., 18 (2018), 347–369. http://dx.doi.org/10.1007/s12346-018-0289-9 doi: 10.1007/s12346-018-0289-9
|
| [31] | G. Wang, M. Yi, S. Tang, Dynamics of an antitumour model with pulsed radioimmunotherapy, Comput. Math. Methods Med., 2022. http://dx.doi.org/https://doi.org/10.1155/2022/4692772 |
| [32] |
W. Wang, X. Lai, Global stability analysis of a viral infection model in a critical case, Math. Biosci. Eng., 17 (2020), 1442–1449. http://dx.doi.org/10.3934/mbe.2020074 doi: 10.3934/mbe.2020074
|
| [33] |
W. Li, T. Zhang, Y. Wang, H. Cheng, Dynamic analysis of a plankton–herbivore state-dependent impulsive model with action threshold depending on the density and its changing rate, Nonlinear Dynam., 107 (2022), 2951–2963. http://dx.doi.org/10.1007/s11071-021-07022-w doi: 10.1007/s11071-021-07022-w
|
| [34] |
B. Tang, W. Zhao, Sliding dynamics and bifurcations of a filippov system with nonlinear threshold control, Int. J. Bifurc. Chaos, 31 (2021), 2150214. http://dx.doi.org/10.1142/S021812742150214X doi: 10.1142/S021812742150214X
|
| [35] | Y. Tian, H. Li, The study of a predator-prey model with fear effect based on state-dependent harvesting strategy, Complexity, 2022. http://dx.doi.org/10.1155/2022/9496599 |
| [36] |
Q. Zhang, B. Tang, S. Tang, Vaccination threshold size and backward bifurcation of sir model with state-dependent pulse control, J. Theoret. Biol., 455 (2018), 75–85. http://dx.doi.org/10.1016/j.jtbi.2018.07.010 doi: 10.1016/j.jtbi.2018.07.010
|
| [37] |
R. J Smith, E. J. Schwartz, Predicting the potential impact of a cytotoxic t-lymphocyte hiv vaccine: How often should you vaccinate and how strong should the vaccine be?, Math. Biosci., 212 (2008), 180–187. http://dx.doi.org/10.1016/j.mbs.2008.02.001 doi: 10.1016/j.mbs.2008.02.001
|
| [38] |
B. Tang, Y. Xiao, S. Tang, R. A. Cheke, A feedback control model of comprehensive therapy for treating immunogenic tumours, Int. J. Bifurc. Chaos, 26 (2016), 1650039. http://dx.doi.org/10.1142/S0218127416500395 doi: 10.1142/S0218127416500395
|
| [39] |
Q. Zhang, S. Tang, Bifurcation analysis of an ecological model with nonlinear state–dependent feedback control by poincaré map defined in phase set, Commun. Nonlinear Sci. Numer. Simul., 108 (2022), 106212. http://dx.doi.org/https://doi.org/10.1016/j.cnsns.2021.106212 doi: 10.1016/j.cnsns.2021.106212
|
| [40] |
Z. Liang, G. Pang, X. Zeng, Y. Liang, Qualitative analysis of a predator–prey system with mutual interference and impulsive state feedback control, Nonlinear Dynam., 87 (2017), 1495–1509. http://dx.doi.org/10.1007/s11071-016-3129-y doi: 10.1007/s11071-016-3129-y
|
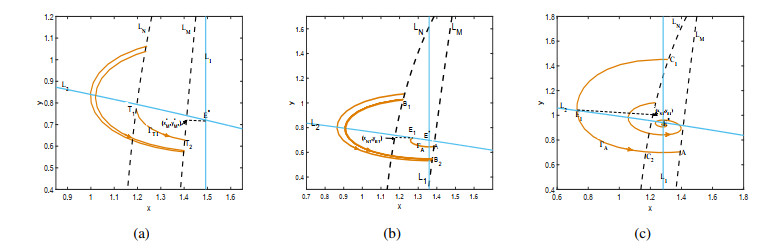
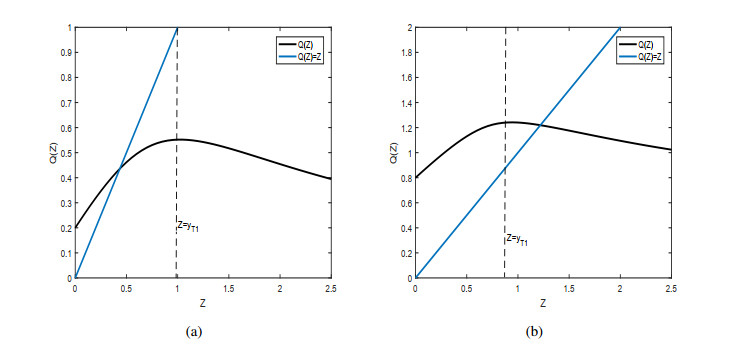
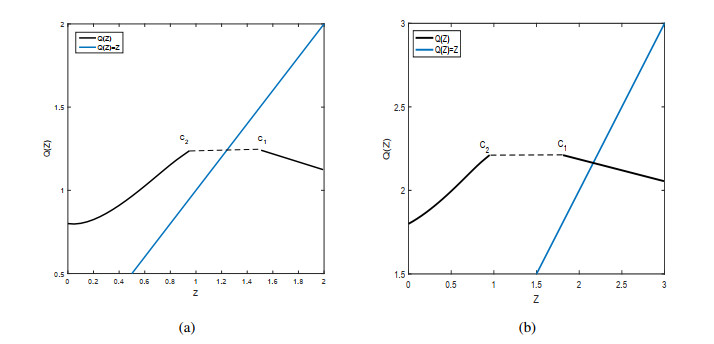
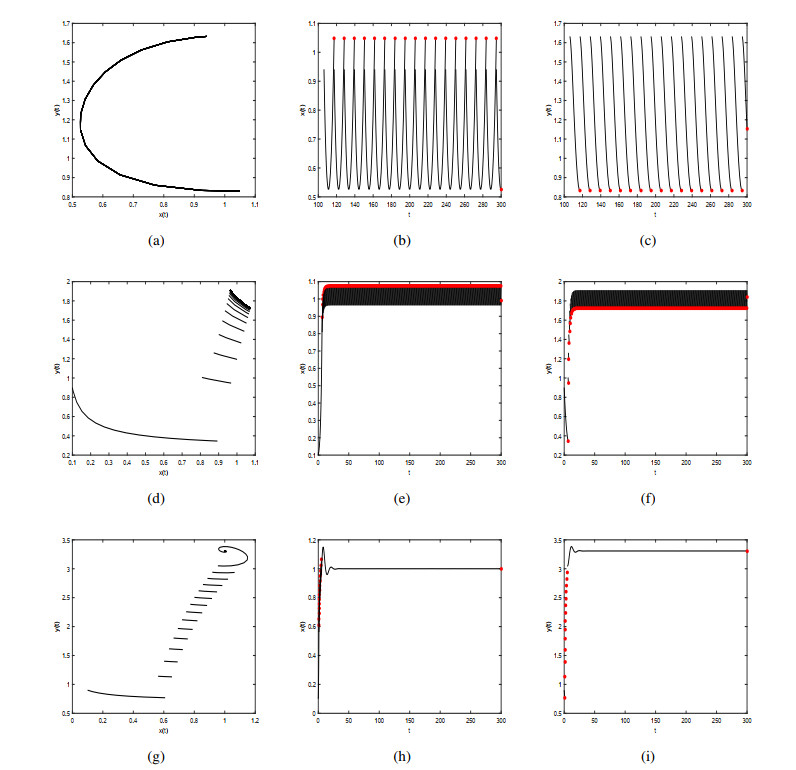
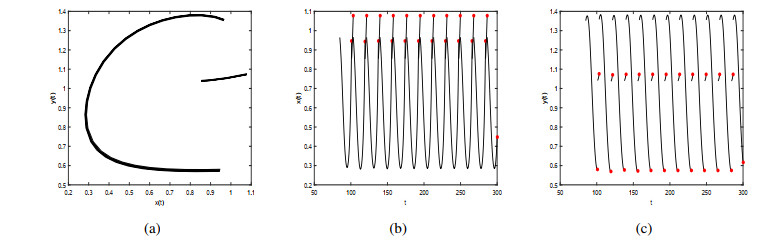
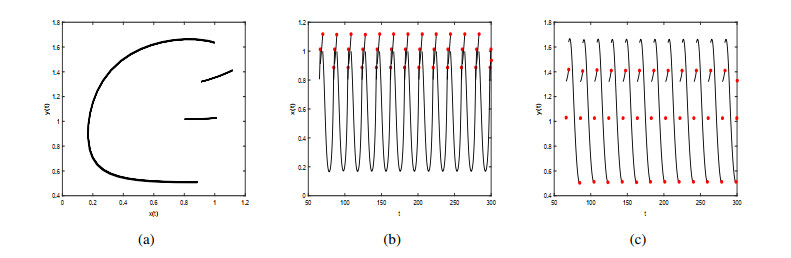
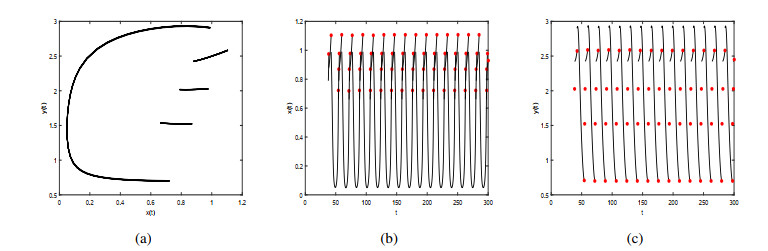
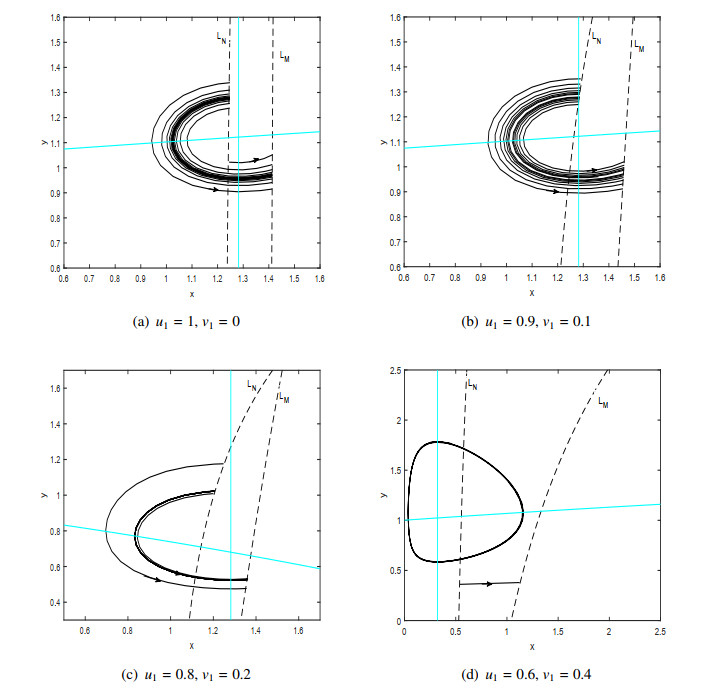
Figures(9)

Yazhi Wu, Guangyao Tang, Changcheng Xiang. Dynamic analysis of a predator-prey state-dependent impulsive model with fear effect in which action threshold depending on the prey density and its changing rate[J]. Mathematical Biosciences and Engineering, 2022, 19(12): 13152-13171. doi: 10.3934/mbe.2022615







 DownLoad:
DownLoad: