Citation: Vladimir A. Kaminsky, Nina Yu. Obvintseva, Svetlana A. Epshtein. The estimation of the kinetic parameters of low-temperature coal oxidation[J]. AIMS Energy, 2017, 5(2): 163-172. doi: 10.3934/energy.2017.2.163

| [1] | Carras JN, Young BC (1994) Self-heating of coal and related materials: Models, application and test methods. Prog Energ Combust 1-15. |
| [2] |
Wang H, Dlugogorski BZ, Kennedy EM (2003) Coal oxidation at low temperatures: Oxygen consumption, oxidation products, reaction mechanism and kinetic modelling. Prog Energ Combust 29: 487-513. doi: 10.1016/S0360-1285(03)00042-X

|
| [3] | Bowes PC (1984) Self-heating: Evaluating and Controlling the Hazards. Amsterdam, The Netherlands: Elsevier Press. |
| [4] |
Beamish BB, Barakat MA, George JDS (2000) Adiabatic testing procedures for determining the self-heating propensity of coal and sample ageing effects. Thermochim Acta 362: 79-87. doi: 10.1016/S0040-6031(00)00588-8
|
| [5] |
Qi X, Xin H, Wang D, et al. (2013) A rapid method for determining the R70 self-heating rate of coal. Thermochim Acta 571: 21-27. doi: 10.1016/j.tca.2013.08.008
|
| [6] |
Nugroho YS, McIntosh AC, Gibbs BM (1998) Using the crossing point method to assess the self-heating behavior of indonesian coals. Symp (International) Combust 27: 2981-2989. doi: 10.1016/S0082-0784(98)80158-6
|
| [7] |
Xuyao Q, Wang D, Milke JA, et al. (2011) Crossing point temperature of coal. Min Sci Technol (China) 21: 255-260. doi: 10.1016/j.mstc.2011.02.024
|
| [8] |
Avila C, Wu T, Lester E (2014) Estimating the Spontaneous Combustion Potential of Coals Using Thermogravimetric Analysis. Energ Fuel 28: 1765-1773. doi: 10.1021/ef402119f
|
| [9] | Li Z, Zhang Y, Jing X, et al. (2015) Insight into the intrinsic reaction of brown coal oxidation at low temperature: Differential scanning calorimetry study. Fuel Process Technol 147: 64-70. |
| [10] | Krishnaswamy S, Bhat S, Gunn RD, et al. (1996) Low-temperature oxidation of coal 1. A single-particle reaction-diffusion model. Fuel 75: 333-343. |
| [11] |
Li B, Chen G, Zhang H, et al. (2014) Development of non-isothermal TGA–DSC for kinetics analysis of low temperature coal oxidation prior to ignition. Fuel 118: 385-391. doi: 10.1016/j.fuel.2013.11.011
|
| [12] |
Chen G, Ma X, Lin M, et al. (2015) Study on thermochemical kinetic characteristics and interaction during low temperature oxidation of blended coals. J Energ Inst 88: 221-228. doi: 10.1016/j.joei.2014.09.007
|
| [13] |
Arisoy A, Beamish B (2015) Reaction kinetics of coal oxidation at low temperatures. Fuel 159: 412-417. doi: 10.1016/j.fuel.2015.06.054
|
| [14] | Nelson MI, Chen XD (2007) Survey of experimental work on the self-heating and spontaneous combustion of coal. Reviews in Engineering Geology:Geol Soc Am 31-83. |
| [15] | Parr SW, Coons CC (1925) Carbon Dioxide as an Index of the Critical Oxidation Temperature for Coal in Storage. Ind Eng Chem 17: 118-120. |
| [16] |
Sujanti W, Zhang DK, Chen XD (1999) Low-temperature oxidation of coal studied using wire-mesh reactors with both steady-state and transient methods. Combust Flame 117: 646-651. doi: 10.1016/S0010-2180(98)00139-4
|
| [17] | Chen X, Chong L (1995) Some Characteristics of Transient Self-Heating Inside an Exothermically Reactive Porous Solid Slab. Process Saf Environ 73: 101-107. |
| [18] |
Chen X, Sidhu H, Nelson M (2013) A linear relationship between dimensionless crossing-point-temperature and Frank–Kamenetskii reactivity parameter in self-heating test at infinite Biot number for slab geometry. Fire Safety J 61: 138-143. doi: 10.1016/j.firesaf.2013.08.002
|
| [19] | Frank-Kamenet︠skii DA (1969) Diffusion and heat transfer in chemical kinetics. New York: Plenum Press. p.370 p. |
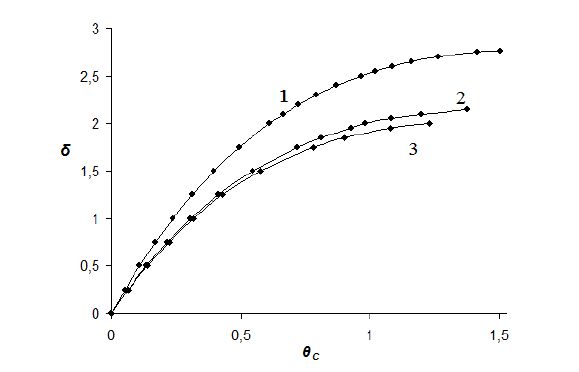
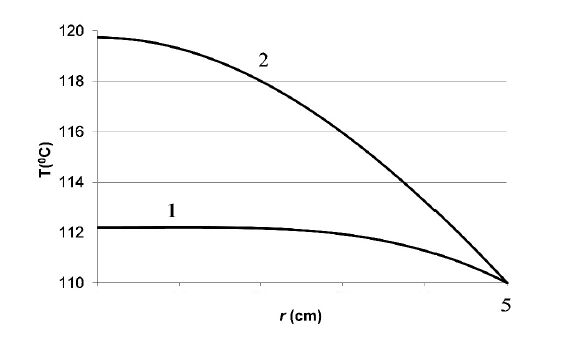
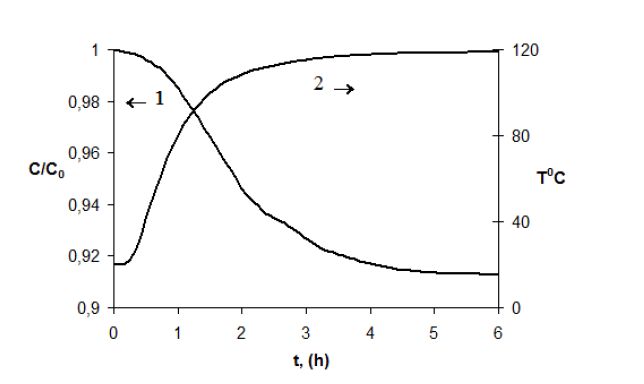
Figures(3) / Tables(1)

Vladimir A. Kaminsky, Nina Yu. Obvintseva, Svetlana A. Epshtein. The estimation of the kinetic parameters of low-temperature coal oxidation[J]. AIMS Energy, 2017, 5(2): 163-172. doi: 10.3934/energy.2017.2.163







 DownLoad:
DownLoad: