Citation: Mikaeel Ahmadi, Mir Sayed Shah Danish, Mohammed Elsayed Lotfy, Atsushi Yona, Ying-Yi Hong, Tomonobu Senjyu. Multi-objective time-variant optimum automatic and fixed type of capacitor bank allocation considering minimization of switching steps[J]. AIMS Energy, 2019, 7(6): 792-818. doi: 10.3934/energy.2019.6.792

| [1] | Size and Location of Capacitor in Electrical System-(Part1). Electrical Notes & Articles (2013). Available from: https://electricalnotes.wordpress.com/2013/02/06/size-and-location-of-capacitorin-electrical-system-part1/. |
| [2] | Venkataramana A, Carr J, Ramshaw RS (1987) Optimal reactive power allocation. IEEE Trans Power Syst 7: 138-144. |
| [3] |
Sundhararajan S, Pahwa A (1994) Optimal selection of capacitors for radial distribution systems using a genetic algorithm. IEEE Trans Power Syst 9: 1499-1507. doi: 10.1109/59.336111

|
| [4] |
Azevedo MSS, Abril IP, Leite JC, et al. (2016) Capacitors placement by NSGA-II in distribution systems with non-linear loads. Int J Electr Power Energy Syst 82: 281-287. doi: 10.1016/j.ijepes.2016.03.025
|
| [5] |
Javadi MS, Esmaeel Nezhad A, Siano P, et al. (2017) Shunt capacitor placement in radial distribution networks considering switching transients decision making approach. Int J Electr Power Energy Syst 92: 167-180. doi: 10.1016/j.ijepes.2017.05.001
|
| [6] |
Chiou JP, Chang CF (2015) Development of a novel algorithm for optimal capacitor placement in distribution systems. Int J Electr Power Energy Syst 73: 684-690. doi: 10.1016/j.ijepes.2015.06.003
|
| [7] |
El-fergany AA, Abdelaziz A (2014) Capacitor allocations in radial distribution networks using cuckoo search algorithm. IET Gener, Transm Distrib 8: 223-232. doi: 10.1049/iet-gtd.2013.0290
|
| [8] |
Abou El-Ela AA, El-Sehiemy RA, Kinawy AM, et al. (2016) Optimal capacitor placement in distribution systems for power loss reduction and voltage improvement. IET Gener, Transm Distrib 10: 1209-1221. doi: 10.1049/iet-gtd.2015.0799
|
| [9] |
Pazouki S, Mohsenzadeh A, Haghifam M, et al. (2015) Simultaneous allocation of charging stations and capacitors in distribution networks improving voltage and power loss. Can J Electr Comput Eng 38: 100-105. doi: 10.1109/CJECE.2014.2377653
|
| [10] |
Yan X, Zhao YD, Kit PW, et al. (2013) Optimal capacitor placement to distribution transformers for power loss reduction in radial distribution systems. IEEE Trans Power Syst 28: 4072-4079. doi: 10.1109/TPWRS.2013.2273502
|
| [11] |
da Rosa WM, Rossoni P, Teixeira JC, et al. (2016) Optimal allocation of capacitor banks using genetic algorithm and sensitivity analysis. IEEE Lat Am Trans 14: 3702-3707. doi: 10.1109/TLA.2016.7786353
|
| [12] |
Askarzadeh A (2016) Capacitor placement in distribution systems for power loss reduction and voltage improvement: A new methodology. IET Gener, Transm Distrib 10: 3631-3638. doi: 10.1049/iet-gtd.2016.0419
|
| [13] |
Das D (2002) Reactive power compensation for radial distribution networks using genetic algorithm. Int J Electr Power Energy Syst 24: 573-581. doi: 10.1016/S0142-0615(01)00068-0
|
| [14] |
Ghanegaonkar S, Pande V (2017) Optimal hourly scheduling of distributed generation and capacitors for minimisation of energy loss and reduction in capacitors switching operations. IET Gener, Transm Distrib 11: 2244-2250. doi: 10.1049/iet-gtd.2016.1600
|
| [15] | Carpinelli G, Mottola F, Proto D, et al. (2010) Optimal allocation of dispersed generators capacitors and distributed energy storage systems in distribution networks. 2010 Modern Electric Power Systems, Wroclaw, Poland, 1-6. |
| [16] |
Rahmani-andebili M (2015) Reliability and economic-driven switchable capacitor placement in distribution network. IET Gener, Transm Distrib 9: 1572-1579. doi: 10.1049/iet-gtd.2015.0359
|
| [17] |
Rahmani-andebili M (2016) Simultaneous placement of DG and capacitor in distribution network. Electr power syst res 131: 1-10. doi: 10.1016/j.epsr.2015.09.014
|
| [18] | Mohagheghi E, Alramlawi M, Gabash A, et al. (2018) A survey of Real-Time optimal power flow. Energies 11: 1-20. |
| [19] | Mohagheghi E, Gabash A, Li Pu (2017) A framework for Real-Time optimal power flow under wind energy penetration. Energies 10: 1-28. |
| [20] | Jizhong Z (2009) Optimization of power system operation. IEEE Wiley Press, John Wiley & Sons, Hoboken, NJ, USA. |
| [21] | Hadi S (2011) Power System Analysis. PSA Publishing LLC, 3rd Eds. |
| [22] |
Rahmani-andebili M (2016) Economic and network-driven distributed generation placement modeling feeder's failure rate and customer's load type. IEEE Trans Ind Electron 63: 1598-1606. doi: 10.1109/TIE.2015.2498902
|
| [23] | Herrero J (2006) Non-linear robust identification using evolutionary algorithms. Doctoral thesis, Polytechnic University. Valencia. |
| [24] |
Laumanns M, Thiele L, Deb K, et al. (2002) Combining convergence and diversity in evolutionary multi-objective optimization. Evol comput 10: 263-282. doi: 10.1162/106365602760234108
|
| [25] | Ahmadi M, Lotfy M, Shigenobu R, et al. (2019) Optimal sizing of multiple renewable energy resources and PV inverter reactive power control encompassing environmental, technical, and economic issues. IEEE Syst J 1-12. |
| [26] |
Ahmadi M, Lotfy M, Danish MSS, et al. (2019) Optimal multi-configuration and allocation of SVR, capacitor, centralised wind farm, and energy storage system: A multi-objective approach in a real distribution network. IET Renewable Power Gener 13: 762-773. doi: 10.1049/iet-rpg.2018.5057
|
| [27] | Herrero J, Meza G, Martinez M, et al. (2014) A smart-distributed pareto front using the ev-MOGA evolutionary algorithm. Int J Artif Intell Tools 23: 145002-1-145002-22. |
| [28] | Herrero J, Blasco X, Martinez M, et al. (2008) Multiobjective tuning of robust PID controllers using evolutionary algorithms. Lect Notes Comput Sci 515-524. |
| [29] |
Herrero J, Blasco X, Sanchis J, et al. (2016) Design of sound phase diffusers by means of multiobjective optimization approach using ev-MOGA evolutionary algorithm. Struct Multidiscip Optim 53: 861-879. doi: 10.1007/s00158-015-1367-0
|
| [30] |
Ahmadi M, Lotfy ME, Shigenobu R, et al. (2018) Optimal sizing and placement of rooftop solar photovoltaic at Kabul city real distribution network. IET Gener, Transm Distrib 12: 303-309. doi: 10.1049/iet-gtd.2017.0687
|
| [31] |
Ahmadi M, Lotfy M, Howlader A, et al. (2019) Centralized multi-objective integration of wind farm and battery energy storage system in real distribution network considering environmental, technical and economic perspective. IET Gener, Transm Distrib 13: 5207-5217. doi: 10.1049/iet-gtd.2018.6749
|
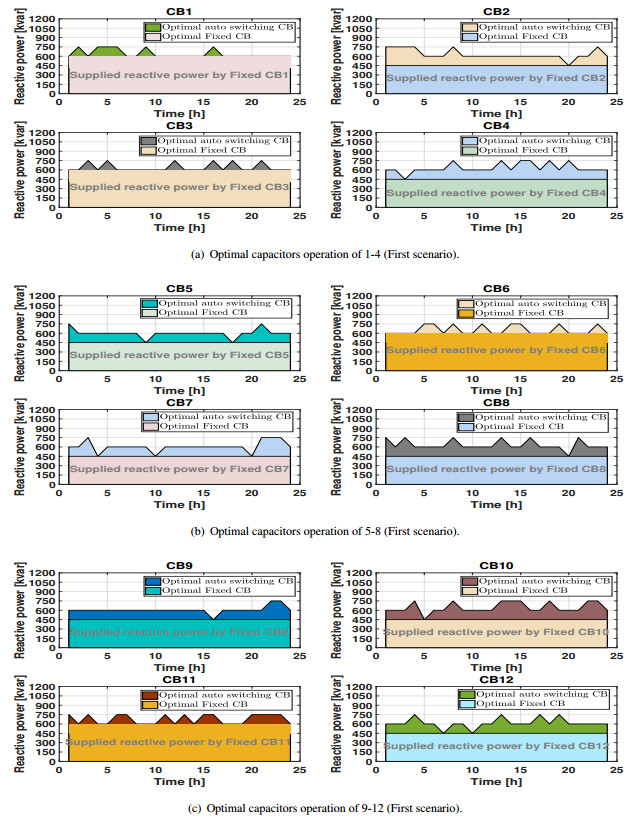
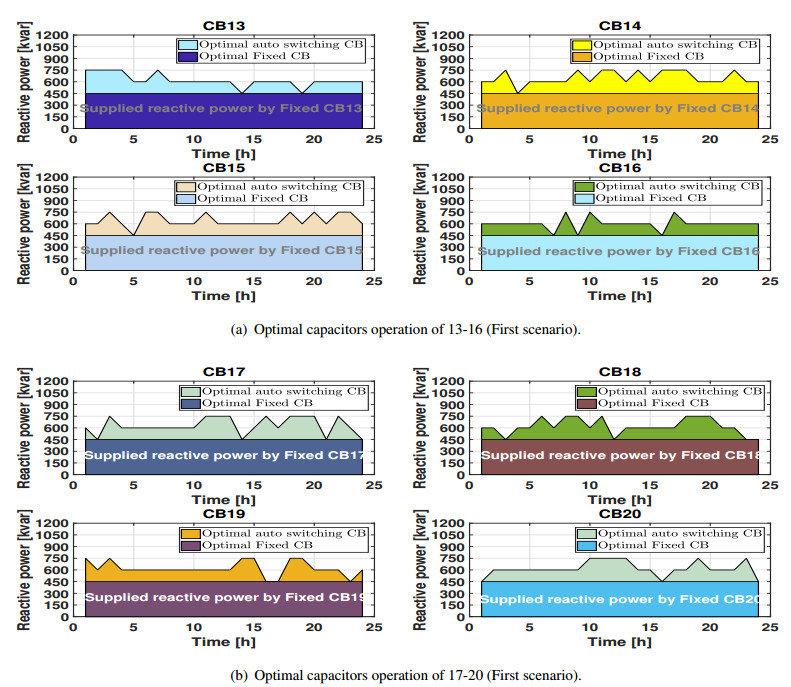
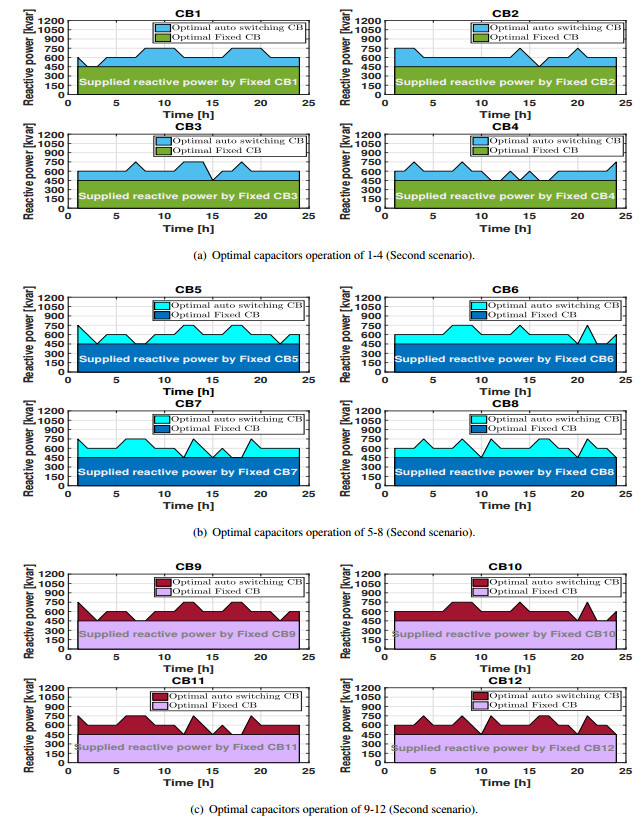
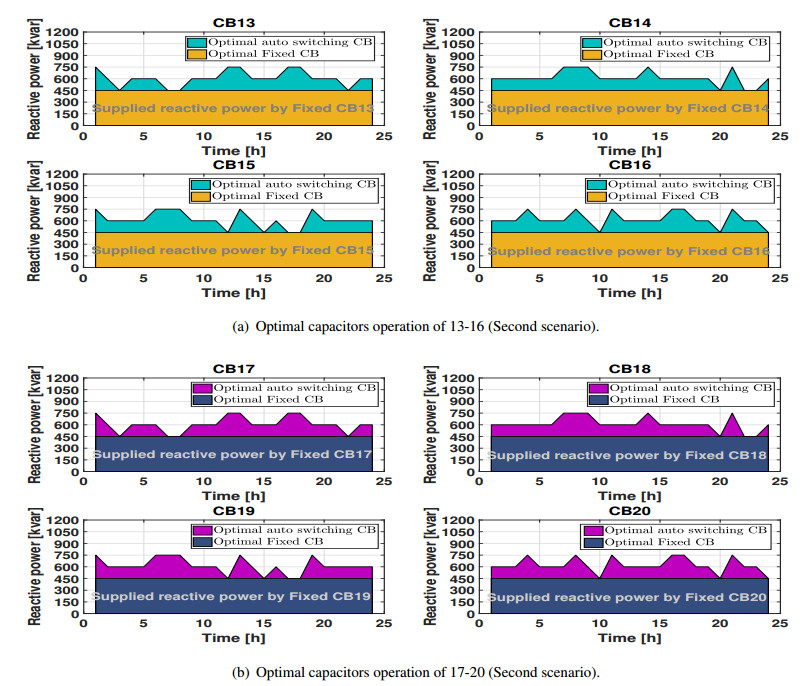
Figures(11) / Tables(3)

Mikaeel Ahmadi, Mir Sayed Shah Danish, Mohammed Elsayed Lotfy, Atsushi Yona, Ying-Yi Hong, Tomonobu Senjyu. Multi-objective time-variant optimum automatic and fixed type of capacitor bank allocation considering minimization of switching steps[J]. AIMS Energy, 2019, 7(6): 792-818. doi: 10.3934/energy.2019.6.792







 DownLoad:
DownLoad: