Citation: Davood Norouzi, Ataur Katebi, Feng Cui, Victor B. Zhurkin. Topological diversity of chromatin fibers: Interplay between nucleosome repeat length, DNA linking number and the level of transcription[J]. AIMS Biophysics, 2015, 2(4): 613-629. doi: 10.3934/biophy.2015.4.613

| [1] |
van Holde K, Zlatanova J (2007) Chromatin fiber structure: Where is the problem now? Semin Cell Dev Biol 18: 651-658. doi: 10.1016/j.semcdb.2007.08.005

|
| [2] |
Tremethick DJ (2007) Higher-order structures of chromatin: the elusive 30 nm fiber. Cell 128: 651-654. doi: 10.1016/j.cell.2007.02.008
|
| [3] |
Schlick T, Hayes J, Grigoryev S (2012) Toward convergence of experimental studies and theoretical modeling of the chromatin fiber. J Biol Chem 287: 5183-5191. doi: 10.1074/jbc.R111.305763
|
| [4] |
Luger K, Dechassa ML, Tremethick DJ (2012) New insights into nucleosome and chromatin structure an ordered state or a disordered affair? Nat Rev Mol Cell Biol 13: 436-447. doi: 10.1038/nrm3382
|
| [5] | Davey CA, Sargent DF, Luger K, et al. (2002) Solvent mediated interactions in the structure of the nucleosome core particle at 1.9 Å resolution. J Mol Biol 319: 1097-1113. |
| [6] |
Dorigo B, Schalch T, Kulangara A, et al. (2004) Nucleosome arrays reveal the two-start organization of the chromatin fiber. Science 306: 1571-1573. doi: 10.1126/science.1103124
|
| [7] |
Schalch T, Duda S, Sargent DF, et al. (2005) X-ray structure of a tetra-nucleosome and its implications for the chromatin fiber. Nature 436: 138-141. doi: 10.1038/nature03686
|
| [8] |
Song F, Chen P, Sun D, et al. (2014) Cryo-EM study of the chromatin fiber reveals a double helix twisted by tetranucleosomal units. Science 344: 376-380. doi: 10.1126/science.1251413
|
| [9] |
Routh A, Sandin S, Rhodes D (2008) Nucleosome repeat length and linker histone stoichiometry determine chromatin fiber structure. Proc Natl Acad Sci U S A 105: 8872-8877. doi: 10.1073/pnas.0802336105
|
| [10] | Robinson PJJ, Fairall L, Huynh VAT, et al. (2006) EM measurements define the dimensions of the “30-nm” chromatin fiber: evidence for a compact, interdigitated structure. Proc Natl Acad Sci U S A 103: 6506-6511. |
| [11] |
Lowary PT, Widom J (1998) New DNA sequence rules for high affinity binding to histone octamer and sequence-directed nucleosome positioning. J Mol Biol 276: 19-42. doi: 10.1006/jmbi.1997.1494
|
| [12] |
Lohr DE (1981) Detailed analysis of the nucleosomal organization of transcribed DNA in yeast chromatin. Biochemistry, 20: 5966-5972. doi: 10.1021/bi00524a007
|
| [13] | Strauss F, Prunell A (1983) Organization of internucleosomal DNA in rat liver chromatin. EMBO J 2: 51-56. |
| [14] |
Wang J, Fondufe-Mittendorf Y, Xi L, et al. (2008) Preferentially quantized linker DNA lengths in Saccharomyces cerevisiae. PLoS Comput Biol 4: e1000175. doi: 10.1371/journal.pcbi.1000175
|
| [15] |
Cui F, Cole HA, Clark DJ, et al. (2012) Transcriptional activation of yeast genes disrupts intragenic nucleosome phasing. Nucleic Acids Res 40: 10753-10764. doi: 10.1093/nar/gks870
|
| [16] |
Correll SJ, Schubert MH, Grigoryev SA (2012) Short nucleosome repeats impose rotational modulations on chromatin fibre folding. EMBO J 31: 2416-2426. doi: 10.1038/emboj.2012.80
|
| [17] |
Worcel A, Strogatz S, Riley D (1981) Structure of chromatin and the linking number of DNA. Proc Natl Acad Sci U S A 78: 1461-1465. doi: 10.1073/pnas.78.3.1461
|
| [18] |
Woodcock CL, Frado LL, Rattner JB (1984) The higher-order structure of chromatin: evidence for a helical ribbon arrangement. J Cell Biol 99: 42-52. doi: 10.1083/jcb.99.1.42
|
| [19] |
Williams SP, Athey BD, Muglia LJ, et al. (1986) Chromatin fibers are left-handed double helices with diameter and mass per unit length that depend on linker length. Biophys J 49: 233-248. doi: 10.1016/S0006-3495(86)83637-2
|
| [20] |
Fogg JM, Randall GL, Pettitt BM, et al. (2012) Bullied no more: when and how DNA shoves proteins around. Q Rev Biophys 45: 257-299. doi: 10.1017/S0033583512000054
|
| [21] |
Baranello L, Levens D, Gupta A, et al. (2012) The importance of being supercoiled: how DNA mechanics regulate dynamic processes. Biochim Biophys Acta 1819: 632-638. doi: 10.1016/j.bbagrm.2011.12.007
|
| [22] |
Koslover EF, Fuller CJ, Straight AF, et al. (2010) Local geometry and elasticity in compact chromatin structure. Biophys J 99: 3941-3950. doi: 10.1016/j.bpj.2010.10.024
|
| [23] |
Scipioni A, Turchetti G, Morosetti S, et al. (2010) Geometrical, conformational and topological restraints in regular nucleosome compaction in chromatin. Biophys Chem 148: 56-67. doi: 10.1016/j.bpc.2010.02.010
|
| [24] |
Norouzi D, Zhurkin VB (2015) Topological polymorphism of the two-start chromatin fiber. Biophys J 108: 2591-2600. doi: 10.1016/j.bpj.2015.04.015
|
| [25] |
Germond JE, Hirt B, Oudet P, et al. (1975) Folding of the DNA double helix in chromatin-like structures from simian virus 40. Proc Natl Acad Sci U S A 72: 1843-1847. doi: 10.1073/pnas.72.5.1843
|
| [26] |
Simpson RT, Thoma F, Brubaker JM (1985) Chromatin reconstituted from tandemly repeated cloned DNA fragments and core histones: a model system for study of higher order structure. Cell 42: 799-808. doi: 10.1016/0092-8674(85)90276-4
|
| [27] |
Norton VG, Imai BS, Yau P, et al. (1989) Histone acetylation reduces nucleosome core particle linking number change. Cell 57: 449-457. doi: 10.1016/0092-8674(89)90920-3
|
| [28] |
Olson WK, Marky NL, Jernigan RL, et al. (1993) Influence of fluctuations on DNA curvature A comparison of flexible and static wedge models of intrinsically bent DNA. J Mol Biol 232: 530-554. doi: 10.1006/jmbi.1993.1409
|
| [29] | Dickerson RE, Bansal M, Calladine CR, et al. (1989) Definitions and nomenclature of nucleic acid structure parameters. EMBO J 8: 1-4. |
| [30] |
Olson WK, Gorin AA, Lu XJ, et al. (1998) DNA sequence-dependent deformability deduced from protein-DNA crystal complexes. Proc Natl Acad Sci U S A 95: 11163-11168. doi: 10.1073/pnas.95.19.11163
|
| [31] |
Fuller FB (1978) Decomposition of the linking number of a closed ribbon: A problem from molecular biology. Proc Natl Acad Sci U S A 75: 3557-3561. doi: 10.1073/pnas.75.8.3557
|
| [32] |
Crick FH (1976) Linking numbers and nucleosomes. Proc Natl Acad Sci U S A 73: 2639-2643. doi: 10.1073/pnas.73.8.2639
|
| [33] |
Prunell A (1998) A topological approach to nucleosome structure and dynamics: the linking number paradox and other issues. Biophys J 74: 2531-2544. doi: 10.1016/S0006-3495(98)77961-5
|
| [34] |
Levitt M (1983) Protein folding by restrained energy minimization and molecular dynamics. J Mol Biol 170: 723-764. doi: 10.1016/S0022-2836(83)80129-6
|
| [35] | Klenin K, Langowski J (2000) Computation of writhe in modeling of supercoiled DNA. Biopolymers 54: 307-317. |
| [36] |
Zhurkin VB, Lysov YP, Ivanov VI (1979) Anisotropic flexibility of DNA and the nucleosomal structure. Nucleic Acids Res 6: 1081-1096. doi: 10.1093/nar/6.3.1081
|
| [37] |
Ulyanov NB, Zhurkin VB (1984) Sequence-dependent anisotropic flexibility of B-DNA. A conformational study. J Biomol Struct Dyn 2: 361-385. doi: 10.1080/07391102.1984.10507573
|
| [38] |
Gorin AA, Zhurkin VB, Olson WK (1995) B-DNA twisting correlates with base pair morphology. J Mol Biol 247: 34-48. doi: 10.1006/jmbi.1994.0120
|
| [39] |
Lu X-J, Olson WK (2003) 3DNA: A software package for the analysis, rebuilding, and visualization of three-dimensional nucleic acid structures. Nucleic Acids Res 31: 5108-5121. doi: 10.1093/nar/gkg680
|
| [40] | Lu X-J, El Hassan MA, Hunter CA (1997) Structure and Conformation of Helical Nucleic Acids: Analysis Program (SCHNAaP) J Mol Biol 273: 668-680. |
| [41] |
Kruithof M, Chien F, Routh A, et al. (2009) Single-molecule force spectroscopy reveals a highly compliant helical folding for the 30-nm chromatin fiber. Nat Struct Mol Biol 16: 534-540. doi: 10.1038/nsmb.1590
|
| [42] |
Keller W (1975) Determination of the number of superhelical turns in simian virus 40 DNA by gel electrophoresis. Proc Natl Acad Sci U S A 72: 4876-4880. doi: 10.1073/pnas.72.12.4876
|
| [43] |
Griffith JD (1975) Chromatin structure: deduced from a minichromosome. Science 187: 1202-1203. doi: 10.1126/science.187.4182.1202
|
| [44] |
Liu LF, Wang JC (1987) Supercoiling of the DNA template during transcription. Proc Natl Acad Sci U S A 84: 7024-7027. doi: 10.1073/pnas.84.20.7024
|
| [45] | Teves SS, Henikoff S (2014) Transcription-generated torsional stress destabilizes nucleosomes. Nat Struct Mol Biol 21: 88-94. |
| [46] |
Cole HA, Howard BH, Clark DJ (2011) The centromeric nucleosome of budding yeast is perfectly positioned and covers the entire centromere. Proc Natl Acad Sci U S A 108: 12687-12692. doi: 10.1073/pnas.1104978108
|
| [47] |
Waern K, Snyder M (2013) Extensive transcript diversity and novel upstream open reading frame regulation in yeast. G3 (Bethesda) 3: 343-352. doi: 10.1534/g3.112.003640
|
| [48] |
Holstege FC, Jennings EG, Wyrick JJ, et al. (1998) Dissecting the regulatory circuitry of a eukaryotic genome. Cell 95: 717-728. doi: 10.1016/S0092-8674(00)81641-4
|
| [49] |
O'Sullivan JM, Tan-Wong SM, Morillon A, et al. (2004) Gene loops juxtapose promoters and terminators in yeast. Nat Genet 36: 1014-1018. doi: 10.1038/ng1411
|
| [50] |
Ansari A, Hampsey M (2005) A role for the CPF 3'-end processing machinery in RNAP II dependent gene looping. Genes Dev 19: 2969-2978. doi: 10.1101/gad.1362305
|
| [51] |
Cole HA, Howard BH, Clark DJ (2011) Activation-induced disruption of nucleosome position clusters on the coding regions of Gcn4-dependent genes extends into neighbouring genes. Nucleic Acids Res 39: 9521-9535. doi: 10.1093/nar/gkr643
|
| [52] |
Hsieh TS, Weiner A, Lajoie B, et al. (2015) Mapping Nucleosome Resolution Chromosome Folding in Yeast by Micro-C. Cell 162: 108-119. doi: 10.1016/j.cell.2015.05.048
|
| [53] |
Teif VB, Vainshtein Y, Caudron-Herger M, et al. (2012) Genome-wide nucleosome positioning during embryonic stem cell development. Nat Struct Mol Biol 19: 1185-1192. doi: 10.1038/nsmb.2419
|
| [54] |
Woodcock CL, Skoultchi AI, Fan Y (2006) Role of linker histone in chromatin structure and function: H1 stoichiometry and nucleosome repeat length. Chromosome Res. 14: 17-25. doi: 10.1007/s10577-005-1024-3
|
| [55] |
Beshnova DA, Cherstvy AG, Vainshtein Y, et al. (2014) Regulation of the nucleosome repeat length in vivo by the DNA sequence, protein concentrations and long-range interactions. PLoS Comput Biol 10: e1003698. doi: 10.1371/journal.pcbi.1003698
|
| [56] |
Cherstvy AG, Teif VB (2014) Electrostatic effect of H1-histone protein binding on nucleosome repeat length. Phys Biol 11: 044001. doi: 10.1088/1478-3975/11/4/044001
|
| [57] |
Kireeva ML, Walter W, Tchernajenko V, et al. (2002) Nucleosome remodeling induced by RNA polymerase II: loss of the H2A/H2B dimer during transcription. Mol Cell 9: 541-552. doi: 10.1016/S1097-2765(02)00472-0
|
| [58] |
Zlatanova J, Victor J (2009) How are nucleosomes disrupted during transcription elongation?. HFSP J 3: 373-378. doi: 10.2976/1.3249971
|
| [59] |
Becavin C, Barbi M, Victor J, et al. (2010) Transcription within condensed chromatin: Steric hindrance facilitates elongation. Biophys J 98: 824-833. doi: 10.1016/j.bpj.2009.10.054
|
| [60] |
Teves SS, Henikoff S (2014) DNA torsion as a feedback mediator of transcription and chromatin dynamics. Nucleus 5: 211-218. doi: 10.4161/nucl.29086
|
| [61] |
Bi X, Broach JR (1997) DNA in transcriptionally silent chromatin assumes a distinct topology that is sensitive to cell cycle progression. Mol Cell Bio 17: 7077-7087. doi: 10.1128/MCB.17.12.7077
|
| [62] |
Tolstorukov MY, Colasanti AV, McCandlish DM, et al. (2007) A novel roll-and-slide mechanism of DNA folding in chromatin: implications for nucleosome positioning. J Mol Biol 371: 725-738. doi: 10.1016/j.jmb.2007.05.048
|
| [63] |
Humphrey W, Dalke A, Schulten K (1996) VMD, Visual molecular dynamics. J Mol Graph 14: 33-8, 27-8. doi: 10.1016/0263-7855(96)00018-5
|
| [64] | Venters BJ, Pugh BF (2009) How eukaryotic genes are transcribed. Crit Rev Biochem Mol Biol 44: 117-141. |
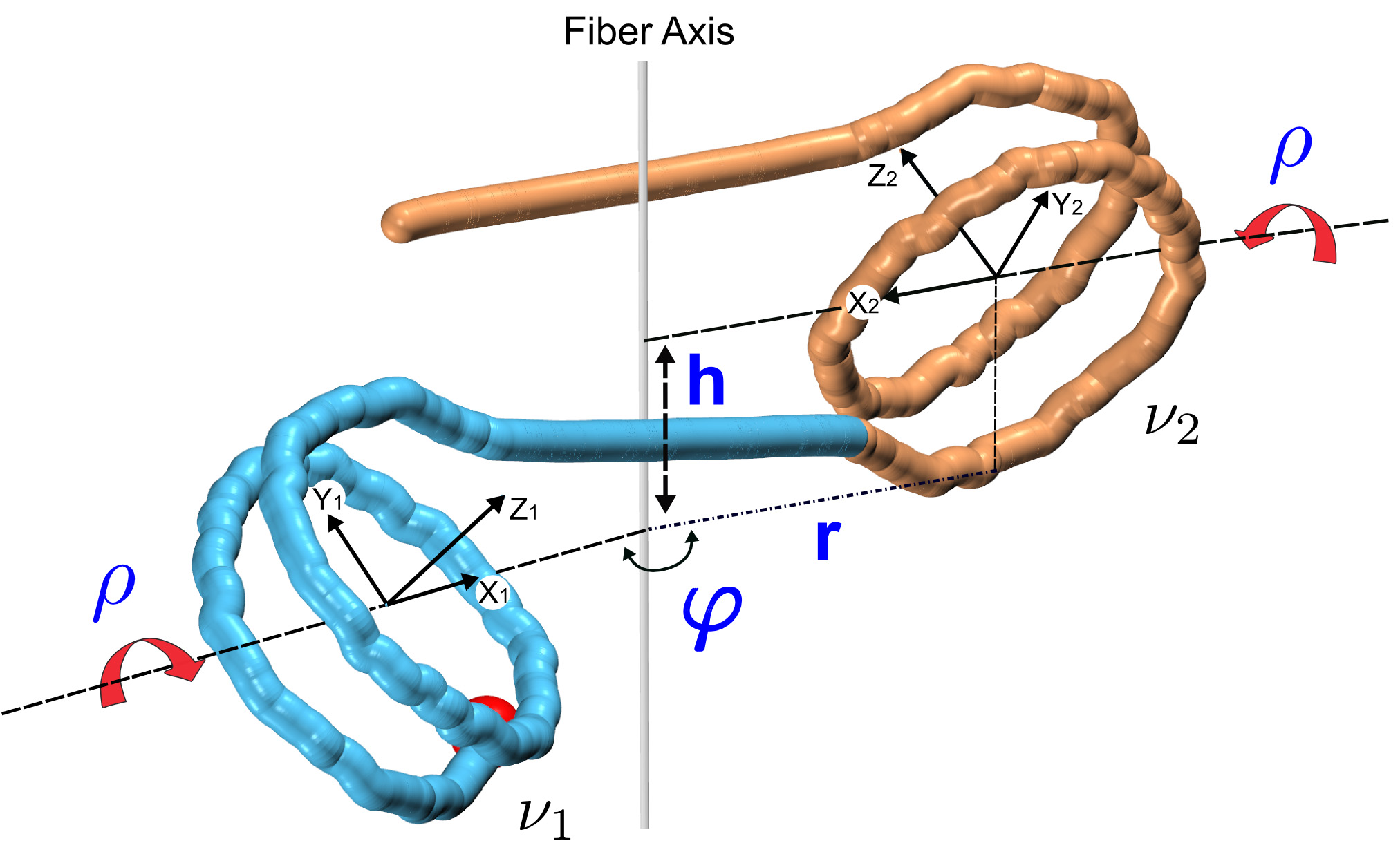
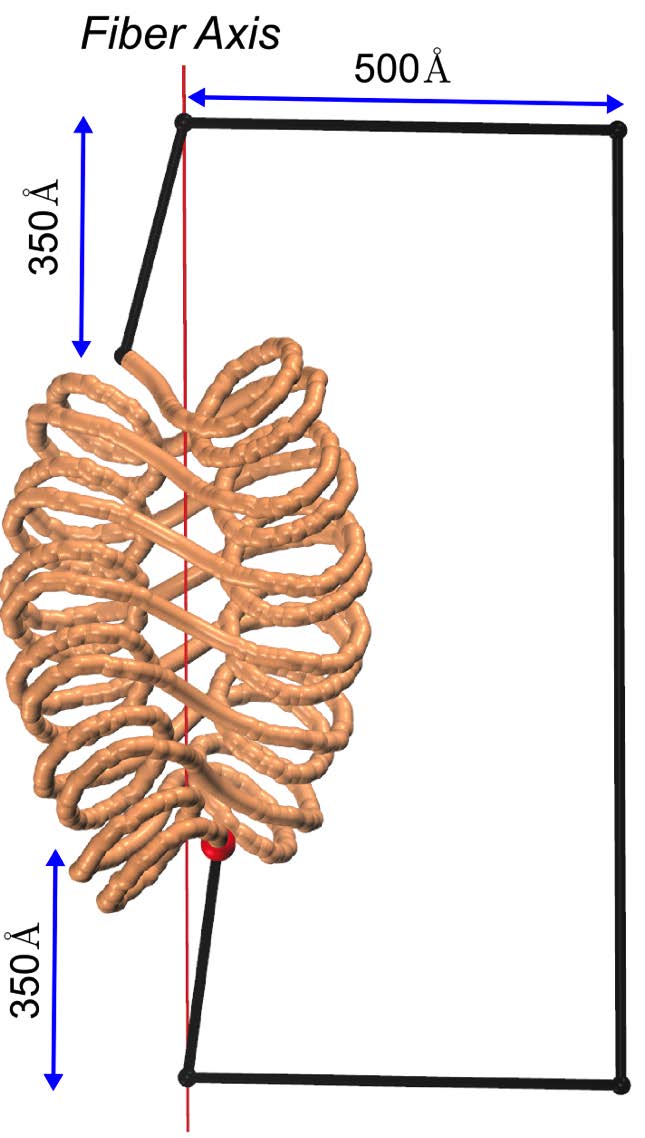
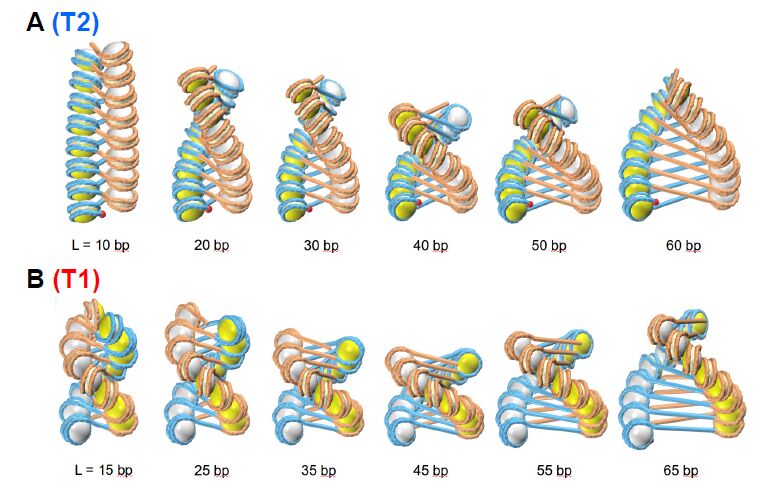
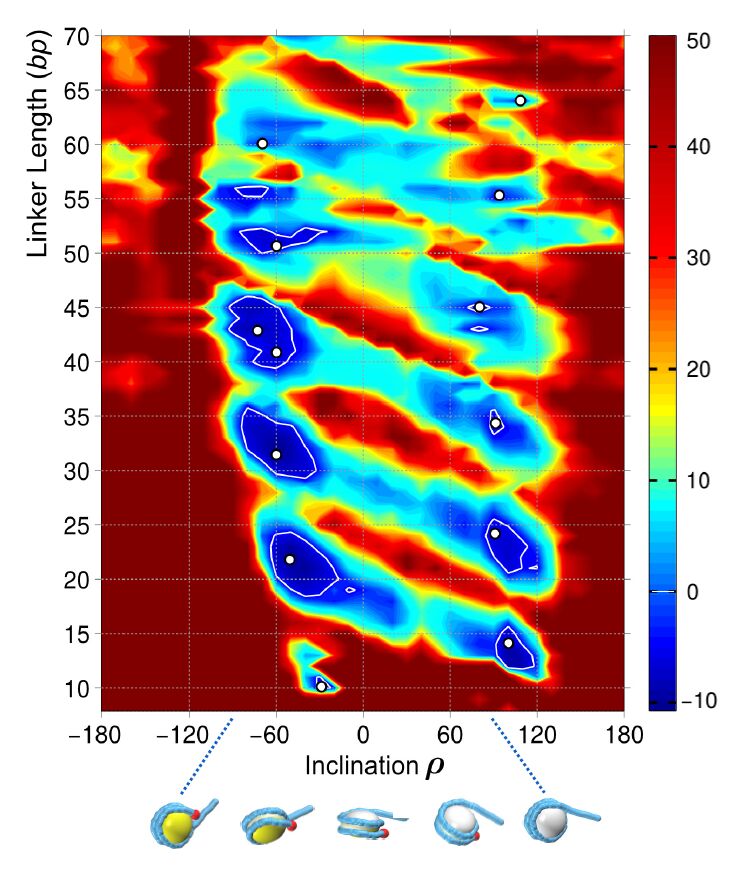
Figures(7)

Davood Norouzi, Ataur Katebi, Feng Cui, Victor B. Zhurkin. Topological diversity of chromatin fibers: Interplay between nucleosome repeat length, DNA linking number and the level of transcription[J]. AIMS Biophysics, 2015, 2(4): 613-629. doi: 10.3934/biophy.2015.4.613







 DownLoad:
DownLoad: