Citation: Ángeles Hidalgo, Francisco-Javier López-Baena, José-Enrique Ruiz-Sainz, José-María Vinardell. Studies of rhizobial competitiveness for nodulation in soybean using a non-destructive split-root system[J]. AIMS Microbiology, 2017, 3(2): 323-334. doi: 10.3934/microbiol.2017.2.323

| [1] | Peix A, Ramírez-Bahena MH, Velázquez E, et al. (2015) Bacterial associations with legumes. Crit Rev Plant Sci 34: 7–42. |
| [2] |
Oldroyd GE (2013) Speak, friend, and enter: signalling systems that promote beneficial symbiotic associations in plants. Nat Rev Microbiol 11: 252–263. doi: 10.1038/nrmicro2990

|
| [3] |
Suzaki T, Kawaguchi M (2014) Root nodulation: a developmental program involving cell fate conversion triggered by symbiotic bacterial infection. Curr Opin Plant Biol 21: 16–22. doi: 10.1016/j.pbi.2014.06.002
|
| [4] | López-Baena FJ, Ruiz-Sainz JE, Rodríguez-Carvajal MA, et al. (2016) Bacterial molecular signals in the Sinorhizobium fredii-soybean symbiosis. Int J Mol Sci 17: E755. |
| [5] |
Graham PH, Vance CP (2000) Nitrogen fixation in perspective: an overview of research and extension needs. Field Crops Res 65: 93–106. doi: 10.1016/S0378-4290(99)00080-5
|
| [6] |
Graham PH, Vance CP (2003) Legumes: importance and constraints to greater use. Plant Physiol 131: 872–877. doi: 10.1104/pp.017004
|
| [7] |
Lindström K, Murwira M, Willems A, et al. (2010) The biodiversity of beneficial microbe-host mutualism: the case of rhizobia. Res Microbiol 161: 453–463. doi: 10.1016/j.resmic.2010.05.005
|
| [8] | Rodríguez-Navarro DN, Margaret-Oliver I, Albareda-Contreras M, et al. (2011) Soybean interactions with soil microbes, agronomical and molecular aspects. Agronomy Sust Developm 31: 173–190. |
| [9] |
Curatti L, Rubio LM (2014) Challenges to develop nitrogen-fixing cereals by direct nif-gene transfer. Plant Sci 225: 130–137. doi: 10.1016/j.plantsci.2014.06.003
|
| [10] |
Dowling DN, Broughton WJ (1986) Competition for nodulation of legumes. Annu Rev Microbiol 40: 131–157. doi: 10.1146/annurev.mi.40.100186.001023
|
| [11] |
Triplett EW, Sadowsky MJ (1992) Genetics of competition for nodulation of legumes. Annu Rev Microbiol 46: 399–428. doi: 10.1146/annurev.mi.46.100192.002151
|
| [12] |
Friesen ML (2012) Widespread fitness alignment in the legume-rhizobium symbiosis. New Phytol 194: 1096–1111. doi: 10.1111/j.1469-8137.2012.04099.x
|
| [13] | Ruiz-Sainz JE, Zhou JC, Rodriguez-Navarro DN, et al. (2005) Soybean cultivation and BNF in China, In: Werner, D., Newton, W.E., Editors., Nitrogen fixation in agriculture, forestry, ecology, and the environment, Dordrecht: Springer, 67–87. |
| [14] |
Thilakarathna MS, Raizada MN (2017) A meta-analysis of the effectiveness of diverse rhizobia inoculants on soybean traits under field conditions. Soil Biol Biochem 105: 177–196. doi: 10.1016/j.soilbio.2016.11.022
|
| [15] |
Delamuta JR, Ribeiro RA, Ormeño-Orrillo E, et al. (2013) Polyphasic evidence supporting the reclassification of Bradyrhizobium japonicum group Ia strains as Bradyrhizobium diazoefficiens sp. nov. Int J Syst Evol Microbiol 63: 3342–3351. doi: 10.1099/ijs.0.049130-0
|
| [16] | Buendía-Clavería AM, Rodríguez-Navarro DN, Santamaría-Linaza C, et al. (1994) Evaluation of the symbiotic properties of Rhizobium fredii in European Soils. Syst Appl Microbiol 17: 155–160. |
| [17] |
Yang SS, Bellogín RA, Buendía A, et al. (2001) Effect of pH and soybean cultivars on the quantitative analyses of soybean rhizobia populations. J Biotechnol 91: 243–255. doi: 10.1016/S0168-1656(01)00340-6
|
| [18] | Pueppke SG, Broughton WJ (1999) Rhizobium sp. strain NGR234 and R. fredii USDA257 share exceptionally broad, nested host ranges. Mol Plant Microbe Interact 12: 293–318. |
| [19] | Annapurna K, Krishnan HB (2003) Molecular aspects of soybean cultivar-specific nodulation by Sinorhizobium fredii USDA257. Indian J Exp Biol 41: 1114–1123. |
| [20] |
Yasuda M, Miwa H, Masuda S, et al. (2016) Effector-triggered immunity determines host genotype-specific incompatibility in legume-Rhizobium symbiosis. Plant Cell Physiol 57: 1791–1800. doi: 10.1093/pcp/pcw104
|
| [21] |
Meinhardt LW, Krishnan HB, Balatti PA, et al. (1993) Molecular cloning and characterization of a sym plasmid locus that regulates cultivar-specific nodulation of soybean by Rhizobium fredii USDA257. Mol Microbiol 9: 17–29. doi: 10.1111/j.1365-2958.1993.tb01665.x
|
| [22] | Rodríguez-Navarro DN, Rodríguez-Carvajal MA, Acosta-Jurado S, et al. (2014) Structure and biological roles of Sinorhizobium fredii HH103 exopolysaccharide. PLoS One 9: e115391. |
| [23] |
Vinardell JM, Ollero FJ, Hidalgo A, et al. (2004) NolR regulates diverse symbiotic signals of Sinorhizobium fredii HH103. Mol Plant-Microbe Interact 17: 676–685. doi: 10.1094/MPMI.2004.17.6.676
|
| [24] |
Vinardell JM, López-Baena FJ, Hidalgo A, et al. (2004) The effect of FITA mutations on the symbiotic properties of Sinorhizobium fredii varies in a chromosomal-background-dependent manner. Arch Microbiol 181: 144–154. doi: 10.1007/s00203-003-0635-3
|
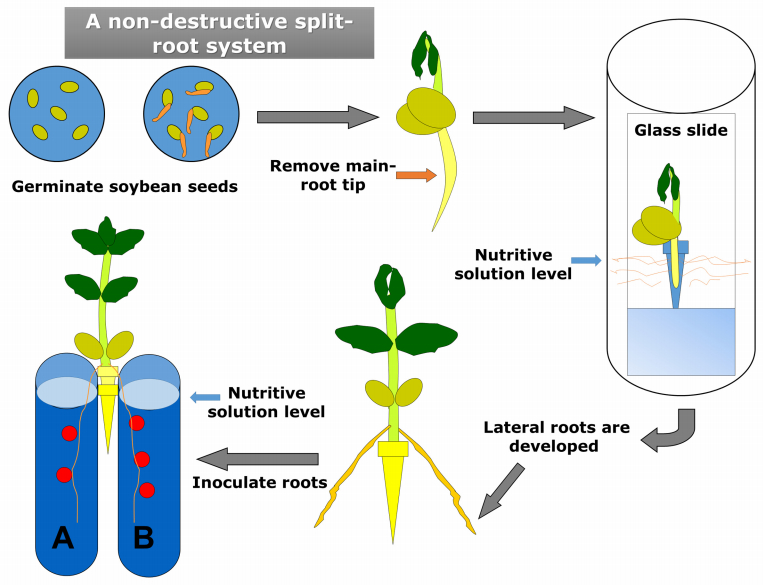
| [25] | Vincent JM (1970) Appendix III. The modified Fåhraeus slide technique, In: Vincent JM, editor, A Manual for the Practical Study of Root Nodule Bacteria, Oxford: Blackwell Scientific, 144–145. |
| [26] | Larrainzar E, Gil-Quintana E, Arrese-Igor C, et al. (2014) Split-root systems applied to the study of the legume-rhizobial symbiosis: what have we learned? J Integr Plant Biol 56: 1118–1124 |
| [27] | Hidalgo A, Ruiz-Sainz JE, Vinardell JM, A new, non-destructive, split-root system for studies with soybean, In: Medina-Morillas C, Lopez Baena J, Editors, Host-Pathogen Interactions: Methods and Protocols, Springer. |
| [28] |
Dowdle SF, Bohlool BB (1987) Intra- and inter-specific competition in Rhizobium fredii and Bradyrhizobium japonicum as indigenous and introduced organisms. Can J Microbiol 33: 990–995. doi: 10.1139/m87-174
|
| [29] |
McLoughlin TJ, Owens PA, Scott A (1985) Competition studies with fast-growing Rhizobium japonicum strains. Can J Microbiol 31: 220–223. doi: 10.1139/m85-042
|
| [30] |
Staehelin C, Krishnan HB (2015) Nodulation outer proteins: double-edged swords of symbiotic rhizobia. Biochem J 470: 263–274. doi: 10.1042/BJ20150518
|
| [31] |
Reid DE, Ferguson BJ, Hayashi S, et al. (2011) Molecular mechanisms controlling legume autoregulation of nodulation. Ann Bot 108: 789–795. doi: 10.1093/aob/mcr205
|
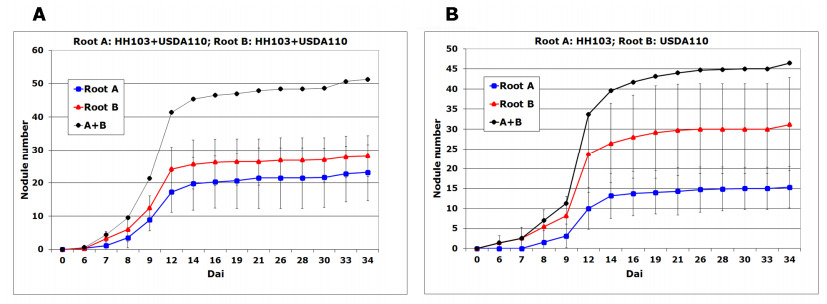
| [32] | Kosslak RM, Bohlool BB (1985) Suppression of nodule development of one side of a split-root system of soybeans caused by prior inoculation of the other side. Plant Physiol 75: 125–130. |
Figures(2) / Tables(2)

Ángeles Hidalgo, Francisco-Javier López-Baena, José-Enrique Ruiz-Sainz, José-María Vinardell. Studies of rhizobial competitiveness for nodulation in soybean using a non-destructive split-root system[J]. AIMS Microbiology, 2017, 3(2): 323-334. doi: 10.3934/microbiol.2017.2.323







 DownLoad:
DownLoad: