Metal matrix composites (MMCs) have been a fundamental element in the development of technologies related to the aerospace and automotive industries. This is because they have an excellent weight-to-strength ratio, i.e., they are light materials with a high mechanical resistance. In the manufacturing of MMCs, the incorporation and homogeneous dispersion of reinforcements in the matrix has been one of the biggest challenges. The issue has expanded to the manufacturing of materials reinforced with nano-scaled particles. This study is aimed at the manufacturing, optimization, and characterization of the polymeric matrix reinforced with carbon nanotubes and alumina (hybrid composites), in order to use the polymeric matrix as an inclusion vehicle of the nano-reinforcements in a metallic matrix.
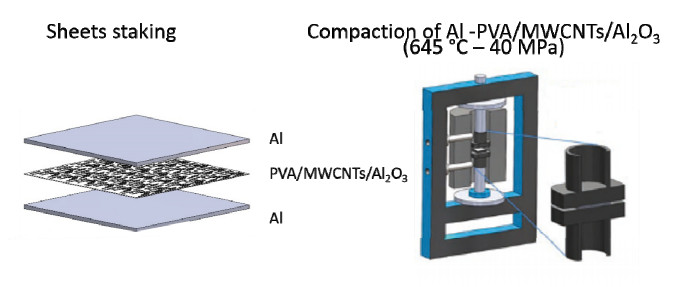
The synthesis of the polymeric matrix composites was carried out by a solution mixing technique using polyvinyl alcohol as a matrix. For the reinforcement's dispersion, a magnetic stirring and sonication were used. Finally, the solution was put into a petri dish to allow its polymerization. The nano-reinforcement dispersion qualification and the quantification of the polymer matrix composite were carried out through the tension, nanoindentation, dynamical mechanical analysis test, elastic modulus mapping, and statistical model for dispersion. In addition, a preliminary study of the metallic composite was carried out and was fabricated by the sandwich technique. The initial characterization of the composites was performed through the nanoindentation test.
Citation: Carlos A. Sánchez, Yamile Cardona-Maya, Andrés D. Morales, Juan S. Rudas, Cesar A. Isaza. Development and evaluation of polyvinyl alcohol films reinforced with carbon nanotubes and alumina for manufacturing hybrid metal matrix composites by the sandwich technique[J]. AIMS Materials Science, 2021, 8(2): 149-165. doi: 10.3934/matersci.2021011
Metal matrix composites (MMCs) have been a fundamental element in the development of technologies related to the aerospace and automotive industries. This is because they have an excellent weight-to-strength ratio, i.e., they are light materials with a high mechanical resistance. In the manufacturing of MMCs, the incorporation and homogeneous dispersion of reinforcements in the matrix has been one of the biggest challenges. The issue has expanded to the manufacturing of materials reinforced with nano-scaled particles. This study is aimed at the manufacturing, optimization, and characterization of the polymeric matrix reinforced with carbon nanotubes and alumina (hybrid composites), in order to use the polymeric matrix as an inclusion vehicle of the nano-reinforcements in a metallic matrix.
The synthesis of the polymeric matrix composites was carried out by a solution mixing technique using polyvinyl alcohol as a matrix. For the reinforcement's dispersion, a magnetic stirring and sonication were used. Finally, the solution was put into a petri dish to allow its polymerization. The nano-reinforcement dispersion qualification and the quantification of the polymer matrix composite were carried out through the tension, nanoindentation, dynamical mechanical analysis test, elastic modulus mapping, and statistical model for dispersion. In addition, a preliminary study of the metallic composite was carried out and was fabricated by the sandwich technique. The initial characterization of the composites was performed through the nanoindentation test.

| [1] |
Isaza MCA, Ledezma Sillas JE, Meza JM, et al. (2017) Mechanical properties and interfacial phenomena in aluminum reinforced with carbon nanotubes manufactured by the sandwich technique. J Compos Mater 51: 1619-1629. doi: 10.1177/0021998316658784

|
| [2] |
Friedrich K, Almajid AA (2013) Manufacturing aspects of advanced polymer composites for automotive applications. Appl Compos Mater 20: 107-128. doi: 10.1007/s10443-012-9258-7
|
| [3] |
Kessler MR (2012) Polymer matrix composites: A perspective for a special issue of polymer reviews. Polym Rev 52: 229-233. doi: 10.1080/15583724.2012.708004
|
| [4] |
Isaza C, Sierra G, Meza J (2016) A novel technique for production of metal matrix composites reinforced with carbon nanotubes. J Manuf Sci E-T ASME 138: 024501. doi: 10.1115/1.4030377
|
| [5] | Alsharef JM, Taha MR, Khan TA (2017) Physical dispersion of nanocarbons in composites-A review. Jurnal Teknologi 79. |
| [6] |
Xin F, Li L (2011) Decoration of carbon nanotubes with silver nanoparticles for advanced CNT/polymer nanocomposites. Compos Part A-Appl S 42: 961-967. doi: 10.1016/j.compositesa.2011.03.024
|
| [7] |
LaNasa JA, Torres VM, Hickey RJ (2020) In situ polymerization and polymer grafting to stabilize polymer-functionalized nanoparticles in polymer matrices. J Appl Phys 127: 134701. doi: 10.1063/1.5144212
|
| [8] |
Khodabakhshi F, Simchi A (2017) The role of microstructural features on the electrical resistivity and mechanical properties of powder metallurgy Al-SiC-Al2O3 nanocomposites. Mater Design 130: 26-36. doi: 10.1016/j.matdes.2017.05.047
|
| [9] |
Khandelwal A, Mani K, Srivastava N, et al. (2017) Mechanical behavior of AZ31/Al2O3 magnesium alloy nanocomposites prepared using ultrasound assisted stir casting. Compos Part B-Eng 123: 64-73. doi: 10.1016/j.compositesb.2017.05.007
|
| [10] |
Kawasaki M, Han JK, Lee DH, et al. (2018) Fabrication of nanocomposites through diffusion bonding under high-pressure torsion. J Mate Res 33: 2700-2710. doi: 10.1557/jmr.2018.205
|
| [11] |
Li Q, Yuan X, Xu H, et al. (2019) Microstructure and fracture toughness of in-situ nanocomposite coating by thermal spraying of Ti3AlC2/Cu powder. Ceram Int 45: 13119-13126. doi: 10.1016/j.ceramint.2019.03.246
|
| [12] |
Merino CAI, Sillas JL, Meza J, et al. (2017) Metal matrix composites reinforced with carbon nanotubes by an alternative technique. J Alloys Compd 707: 257-263. doi: 10.1016/j.jallcom.2016.11.348
|
| [13] |
Isaza MCA, Herrera RamƖrez J, Ledezma Sillas J, et al. (2018) Dispersion and alignment quantification of carbon nanotubes in a polyvinyl alcohol matrix. J Compos Mater 52: 1617-1626. doi: 10.1177/0021998317731151
|
| [14] |
Yuan W, Cui J, Xu S (2016) Mechanical properties and interfacial interaction of modified calcium sulfate whisker/poly(vinyl chloride) composites. J Mater Sci Technol 32: 1352-1360. doi: 10.1016/j.jmst.2016.05.016
|
| [15] | Srivatsan T, Lin Y, Chen F, et al. (2018) Synthesis and microstructural development of particulate reinforced metal-matrix composites using the technique of spray atomization and deposition, TMS Annual Meeting & Exhibition, Springer, Cham, 149-182. |
| [16] |
Koli DK, Agnihotri G, Purohit R (2014) A review on properties, behaviour and processing methods for Al-nano Al2O3 composites. Procedia Mater Sci 6: 567-589. doi: 10.1016/j.mspro.2014.07.072
|
| [17] |
Luo H, Dong J, Xu X, et al. (2018) Exploring excellent dispersion of graphene nanosheets in three-dimensional bacterial cellulose for ultra-strong nanocomposite hydrogels. Compos Part A-Appl S 109: 290-297. doi: 10.1016/j.compositesa.2018.03.007
|
| [18] |
Salom C, Prolongo M, Toribio A, et al. (2018) Mechanical properties and adhesive behavior of epoxy-graphene nanocomposites. Int J Adhes Adhes 84: 119-125. doi: 10.1016/j.ijadhadh.2017.12.004
|
| [19] |
Hu X, Ren N, Chao Y, et al. (2017) Highly aligned graphene oxide/poly(vinyl alcohol) nanocomposite fibers with high-strength, antiultraviolet and antibacterial properties. Compos Part A-Appl S 102: 297-304. doi: 10.1016/j.compositesa.2017.08.015
|
| [20] | Karthikeyan P, Babu BG, Sabarinathan C, et al. (2015) Tribological performance of carbon nanotubes-alumina hybrid/epoxy composites. Optoelectron Adv Mater-Rapid Commun 9: 455-459. |
| [21] | Karthikeyan P, Babu B, Siva K, et al. (2016) Experimental investigation on mechanical behavior of carbon nanotubes-Alumina hybrid epoxy nanocomposites. Dig J Nanomater Bios 11: 625-632. |
| [22] |
Dabees S, Kamel BM, Tirth V, et al. (2020) Experimental design of Al2O3/MWCNT/HDPE hybrid nanocomposites for hip joint replacement. Bioengineered 11: 679-692. doi: 10.1080/21655979.2020.1775943
|
| [23] |
Jiang D, Zhang J, Lv Z (2012) Multi-wall carbon nanotubes (MWCNTs)-SiC composites by laminated technology. J Eur Ceram Soc 32: 1419-1425. doi: 10.1016/j.jeurceramsoc.2011.07.035
|
| [24] |
Ahmad I, Ahmed S, Subhani T, et al. (2016) Synergic influence of MWCNTs and SiC nanoparticles on the microstructure and properties of Al2O3 ceramic hybrid nanocomposites. Curr Appl Phys 16: 1649-1658. doi: 10.1016/j.cap.2016.10.009
|
| [25] |
Han D, Mei H, Xiao S, et al. (2018) A review on the processing technologies of carbon nanotube/silicon carbide composites. J Eur Ceram Soc 38: 3695-3708. doi: 10.1016/j.jeurceramsoc.2018.04.033
|
| [26] |
Padmavathi K, Ramakrishnan R (2014) Tribological behaviour of aluminium hybrid metal matrix composite. Procedia Eng 97: 660-667. doi: 10.1016/j.proeng.2014.12.295
|
| [27] |
Kim H, Babu J, Kang C (2013) Fabrication of A356 aluminum alloy matrix composite with CNTs/Al2O3 hybrid reinforcements. Mat Sci Eng A-Struct 573: 92-99. doi: 10.1016/j.msea.2013.02.041
|
| [28] | Paramsothy M, Chan J, Kwok R, et al. (2012) TiC nanoparticle addition to enhance the mechanical response of hybrid magnesium alloy. J Nanotechnol 2012: 401574. |
| [29] |
Wei TZ, Shamsuri SRB, Yee CS, et al. (2013) Effect of sliding velocity on wear behavior of magnesium composite reinforced with SiC and MWCNT. Procedia Eng 68: 703-709. doi: 10.1016/j.proeng.2013.12.242
|
| [30] | Zhou X, Su D, Wu C, et al. (2012) Tensile mechanical properties and strengthening mechanism of hybrid carbon nanotube and silicon carbide nanoparticle-reinforced magnesium alloy composites. J Nanomater 2012: 851862. |
| [31] | Ramirez JMH, Bustamante RP, Merino CAI, et al. (2020) Unconventional Techniques for the Production of Light Alloys and Composites, Springer, Cham. |
| [32] | SIGMA-ALDRICH Products: Poly(vinyl alcohol) (363081), 2021. Available from: https://www.sigmaaldrich.com/. |
| [33] |
Rossell MD, Kuebel C, Ilari G, et al. (2013) Impact of sonication pretreatment on carbon nanotubes: A transmission electron microscopy study. Carbon 61: 404-411. doi: 10.1016/j.carbon.2013.05.024
|
| [34] |
Chen J, Bull S (2006) On the relationship between plastic zone radius and maximum depth during nanoindentation. Surf Coat Tech 201: 4289-4293. doi: 10.1016/j.surfcoat.2006.08.099
|
| [35] |
Luo Z, Koo JH (2007) Quantifying the dispersion of mixture microstructures. J Microsc 225: 118-125. doi: 10.1111/j.1365-2818.2007.01722.x
|
| [36] |
Olayo R, GarcƖa E, GarcƖa-Corichi B, et al. (1998) Poly(vinyl alcohol) as a stabilizer in the suspension polymerization of styrene: The effect of the molecular weight. J Appl Polym Sci 67: 71-77. doi: 10.1002/(SICI)1097-4628(19980103)67:1<71::AID-APP8>3.0.CO;2-L
|
| [37] |
More S, Dhokne R, Moharil S (2018) Structural properties and temperature dependence dielectric properties of PVA-Al2O3 composite thin films. Polym Bull 75: 909-923. doi: 10.1007/s00289-017-2069-0
|
| [38] |
Mallakpour S, Dinari M (2013) Nanocomposites of poly(vinyl alcohol) reinforced with chemically modified Al2O3: Synthesis and characterization. J Macromol Sci B 52: 1651-1661. doi: 10.1080/00222348.2013.789349
|
| [39] |
Fan B, He D, Liu Y, et al. (2017) Influence of thermal treatments on the evolution of conductive paths in carbon nanotube-Al2O3 hybrid reinforced epoxy composites. Langmuir 33: 9680-9686. doi: 10.1021/acs.langmuir.6b03915
|
| [40] |
Li W, Dichiara A, Zha J, et al. (2014) On improvement of mechanical and thermo-mechanical properties of glass fabric/epoxy composites by incorporating CNT-Al2O3 hybrids. Compos Sci Technol 103: 36-43. doi: 10.1016/j.compscitech.2014.08.016
|
| [41] |
Li W, He D, Dang Z, et al. (2014) In situ damage sensing in the glass fabric reinforced epoxy composites containing CNT-Al2O3 hybrids. Compos Science Technol 99: 8-14. doi: 10.1016/j.compscitech.2014.05.005
|
| [42] |
Haslam MD, Raeymaekers B (2013) A composite index to quantify dispersion of carbon nanotubes in polymer-based composite materials. Composites Part B-Eng 55: 16-21. doi: 10.1016/j.compositesb.2013.05.038
|
| [43] |
Luo Z, Koo J (2008) Quantitative study of the dispersion degree in carbon nanofiber/polymer and carbon nanotube/polymer nanocomposites. Mater Lett 62: 3493-3496. doi: 10.1016/j.matlet.2008.03.010
|



Figures(10) / Tables(2)

Carlos A. Sánchez, Yamile Cardona-Maya, Andrés D. Morales, Juan S. Rudas, Cesar A. Isaza. Development and evaluation of polyvinyl alcohol films reinforced with carbon nanotubes and alumina for manufacturing hybrid metal matrix composites by the sandwich technique[J]. AIMS Materials Science, 2021, 8(2): 149-165. doi: 10.3934/matersci.2021011







 DownLoad:
DownLoad: