Citation: Rumana Islam, Charushi Panditharatne, John Schellenberg, Richard Sparling, Nazim Cicek, David B. Levin. Potential of thin stillage as a low-cost nutrient source for direct cellulose fermentation by Clostridium thermocellum[J]. AIMS Energy, 2015, 3(4): 711-727. doi: 10.3934/energy.2015.4.711

| [1] |
Kapdan IK, Kargi F (2006) Bio-hydrogen production from waste materials. Enzyme Microbial Technol 38: 569-582. doi: 10.1016/j.enzmictec.2005.09.015

|
| [2] |
Solomon BD, Barnes JR, Halvorsen KE (2007) Grain and cellulosic ethanol: History, economics, and energy policy. Biomass Bioenerg 31: 416-425. doi: 10.1016/j.biombioe.2007.01.023
|
| [3] |
Lynd LR, van Zyl WH, McBride JE, et al. (2005) Consolidated bioprocessing of cellulosic biomass: an update. Curr Opinion Biotechnol 16: 577-583. doi: 10.1016/j.copbio.2005.08.009
|
| [4] |
Lynd LR, Weimer PJ, Zyl WH Van, et al. (2002) Microbial Cellulose Utilization : Fundamentals and Biotechnology. Microbiol Mol Biol Rev : MMBR 66: 506-577. doi: 10.1128/MMBR.66.3.506-577.2002
|
| [5] |
Rodrigues LR, Teixeira JA, Oliveira R (2006) Low-cost fermentative medium for biosurfactant production by probiotic bacteria. Biochem Eng J 32: 135-142. doi: 10.1016/j.bej.2006.09.012
|
| [6] |
Kadam KL, Newman MM (1997) Development of a low-cost fermentation medium for ethanol production from biomass. Appl Microbiol Biotechnol 47: 625-629. doi: 10.1007/s002530050985
|
| [7] |
Gullón B, Alonso JL, Parajó JC (2008) Experimental evaluation of alternative fermentation media for L-lactic acid production from apple pomace. J Chem Technol Biotechnol 83: 609-617. doi: 10.1002/jctb.1838
|
| [8] |
Qureshi N, Blaschek HP (2001) Recent advances in ABE fermentation: hyper-butanol producing Clostridium beijerinckii BA101. J Industrial Microbiol Biotechnol 27: 287-291. doi: 10.1038/sj.jim.7000114
|
| [9] |
Parekh M, Formanek J, Blaschek HP (1998) Development of a cost-effective glucose-corn steep medium for production of butanol by Clostridium beijerinckii. J Industrial Microbiol Biotechnol 21: 187-191. doi: 10.1038/sj.jim.2900569
|
| [10] | Schill SR, Fertile Fungus. Ethanol Producer Magazine, 2009. Available from: http://www.ethanolproducer.com/articles/5272/fertile-fungus. |
| [11] |
Schaefer SH, Sung S (2008) Retooling the ethanol industry: thermophilic anaerobic digestion of thin stillage for methane production and pollution prevention. Water Environ Res 80: 101-108. doi: 10.2175/106143007X212157
|
| [12] |
Kim Y, Hendrickson R, Mosier NS, et al. (2008) Enzyme hydrolysis and ethanol fermentation of liquid hot water and AFEX pretreated distillers' grains at high-solids loadings. Bioresour Technol 99: 5206-5215. doi: 10.1016/j.biortech.2007.09.031
|
| [13] |
Agler MT, Garcia ML, Lee ES, et al. (2008) Thermophilic anaerobic digestion to increase the net energy balance of corn grain ethanol. Environ Science Technol 42: 6723-6729. doi: 10.1021/es800671a
|
| [14] | Ratanapariyanuch K, Tyler RT, Shim YY, et al. (2012) Biorefinery process for protein extraction from oriental mustard (Brassica juncea (L.) Czern.) using ethanol stillage. AMB Express 2: 5. |
| [15] |
Davis L, Jeon YJ, Svenson C, et al. (2005) Evaluation of wheat stillage for ethanol production by recombinant Zymomonas mobilis. Biomass Bioenerg 29: 49-59. doi: 10.1016/j.biombioe.2005.02.006
|
| [16] | Weekly Price (2015) Summary—Agriculture and Agri-Food Canada (AAFC), available from: http://www.agr.gc.ca/eng/industry-markets-and-trade/statistics-and-market-information/by-product-sector/crops/crops-market-information-canadian-industry/weekly-price-summary. |
| [17] |
Islam R, Özmihçi S, Cicek N, et al. (2013) Enhanced cellulose fermentation and end-product synthesis by Clostridium thermocellum with varied nutrient compositions under carbon-excess conditions. Biomass bioenerg 48: 213-223. doi: 10.1016/j.biombioe.2012.11.010
|
| [18] | Johnson EA, Madia A, Demain AL (1981) Chemically Defined Minimal Medium for Growth of the Anaerobic Cellulolytic Thermophile Clostridium thermocellum. Appl Environ Microbiol 41: 1060-1062. |
| [19] | Daniels L, Belay N, Rajagopal BS (1986) Assimilatory reduction of sulfate and sulfite by methanogenic bacteria. Appl Envir Microbiol 51: 703-709. |
| [20] |
Islam R, Cicek N, Sparling R, et al. (2009) Influence of initial cellulose concentration on the carbon flow distribution during batch fermentation by Clostridium thermocellum ATCC 27405. Appl microbiol biotechnol 82: 141-148. doi: 10.1007/s00253-008-1763-0
|
| [21] | Grant CL, Pramer D (1962) Minor element composition of yeast extract. J bacteriol 84: 869. |
| [22] |
Islam R, Sparling R, Cicek N, et al. (2015) Optimization of Influential Nutrients during Direct Cellulose Fermentation into Hydrogen by Clostridium thermocellum. Int j mol sci 16: 3116-3132. doi: 10.3390/ijms16023116
|
| [23] | Pauss A, Andre G, Perrier M, et al. (1990) Liquid-to-Gas Mass Transfer in Anaerobic Processes: Inevitable Transfer Limitations of Methane and Hydrogen in the Biomethanation Process. Appl environ microbiol 56: 1636-1644. |
| [24] | Blunt W, Taillefer M, Gapes D, et al. (2015) Evaluating supersaturation of in situ produced gases during thermophilic cellobiose fermentation with Clostridium thermocellum. Transactions of the ASABE 58, In press. |
| [25] |
van Niel EW, Claassen PA, Stams AJ (2003) Substrate and product inhibition of hydrogen production by the extreme thermophile, Caldicellulosiruptor saccharolyticus. Biotechnol Bioeng 81: 255-262. doi: 10.1002/bit.10463
|
| [26] | Schröder C, Selig M, Schönheit P (1994) Glucose fermentation to acetate, CO2 and H2 in the anaerobic hyperthermophilic eubacterium Thermotoga maritima: involvement of the Embden-Meyerhof pathway. Arch microbiol 161: 460-470. |
| [27] |
Nataf Y, Yaron S, Stahl F, et al. (2009) Cellodextrin and laminaribiose ABC transporters in Clostridium thermocellum. J bacterial 191: 203-209. doi: 10.1128/JB.01190-08
|
| [28] | Patni N, Alexander J (1971) Utilization of glucose by Clostridium thermocellum: presence of glucokinase and other glycolytic enzymes in cell extracts. J Bacterio 105: 220-225. |
| [29] |
Hernandez P (1982) Transport of D-glucose in Clostridium thermocellum ATCC-27405. J gen appl microbiol 28: 469-477. doi: 10.2323/jgam.28.469
|
| [30] | Gomez R, Hernandez P (1981) Glucose utilization by Clostridium thermocellum. Advances in Biotechnology. Pergamon Press, Toronto. |
| [31] |
Lu Y, Zhang Y-HP, Lynd LR (2006) Enzyme-microbe synergy during cellulose hydrolysis by Clostridium thermocellum. P Natl Acad Sci USA 103: 16165-16169. doi: 10.1073/pnas.0605381103
|
| [32] | Johnson EA, Reese ET, Demain AL (1982) Inhibition of Clostridium thermocellum cellulase by end products of cellulolysis. J Appl Biochem 4: 64-71. |
| [33] |
Mearls EB, Izquierdo JA, Lynd LR (2012) Formation and characterization of non-growth states in Clostridium thermocellum: spores and L-forms. BMC Microbiol 12: 180. doi: 10.1186/1471-2180-12-180
|
| [34] |
Moreau R, Hicks K (2006) A Reinvestigation of the effect of heat pretreatment of corn fiber and corn germ on the levels of extractable tocopherols and tocotrienols. J Agri Food Chem 54: 8093-8102. doi: 10.1021/jf061422g
|
| [35] |
Ahn JH, Sang BI, Um Y (2011) Butanol production from thin stillage using Clostridium pasteurianum. Bioresour Technol 102: 4934-4937. doi: 10.1016/j.biortech.2011.01.046
|
| [36] | Singh V, Moreau RA, Doner LW, et al. (2007) Recovery of Fiber in the Corn Dry-Grind Ethanol Process: A Feedstock for Valuable Coproducts. Cereal Chem 76: 868-872. |
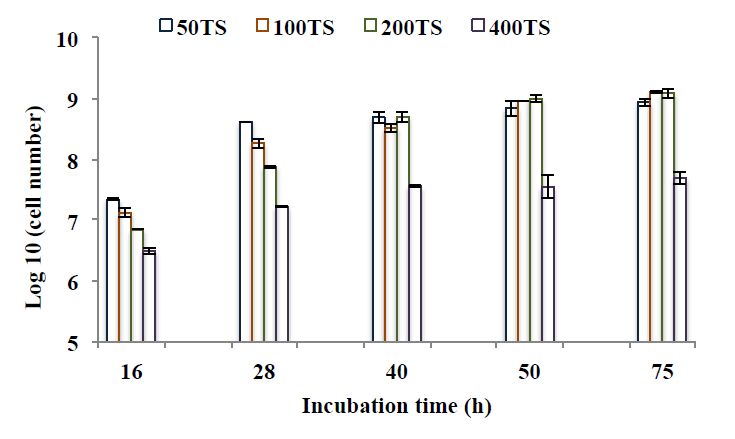
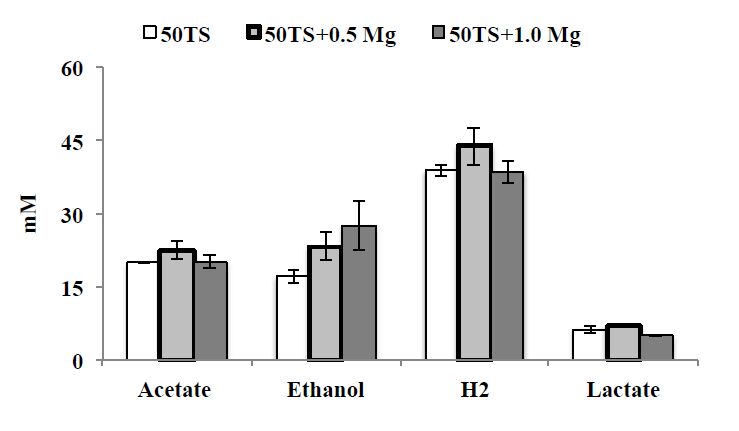
Figures(6) / Tables(3)

Rumana Islam, Charushi Panditharatne, John Schellenberg, Richard Sparling, Nazim Cicek, David B. Levin. Potential of thin stillage as a low-cost nutrient source for direct cellulose fermentation by Clostridium thermocellum[J]. AIMS Energy, 2015, 3(4): 711-727. doi: 10.3934/energy.2015.4.711







 DownLoad:
DownLoad: