Citation: Yoshiji Hantani, Hiroshi Imamura, Tsubasa Yamamoto, Akane Senga, Yuri Yamagami, Minoru Kato, Fusako Kawai, Masayuki Oda. Functional characterizations of polyethylene terephthalate-degrading cutinase-like enzyme Cut190 mutants using bis(2-hydroxyethyl) terephthalate as the model substrate[J]. AIMS Biophysics, 2018, 5(4): 290-302. doi: 10.3934/biophy.2018.4.290

| [1] |
Chen S, Su L, Chen J, et al. (2013) Cutinase: characteristics, preparation, and application. Biotechnol Adv 31: 1754–1767. doi: 10.1016/j.biotechadv.2013.09.005

|
| [2] |
Wei R, Zimmermann W (2017) Biocatalysis as a green route for recycling the recalcitrant plastic polyethylene terephthalate. Microb Biotechnol 10: 1302–1307. doi: 10.1111/1751-7915.12714
|
| [3] | Zimmermann W, Billig S (2010) Enzymes for the biofunctionalization of poly(ethylene terephthalate). Adv Biochem Eng Biotechnol 125: 97–120. |
| [4] |
Kawai F, Oda M, Tamashiro T, et al. (2014) A novel Ca2+-activated, thermostabilized polyesterase capable of hydrolyzing polyethylene terephthalate from Saccharomonospora viridis AHK190. Appl Microbiol Biotech 98: 10053–10064. doi: 10.1007/s00253-014-5860-y
|
| [5] |
Oda M, Yamagami Y, Oida I, et al. (2018) Enzymatic hydrolysis of PET: Functional roles of three Ca2+ ions bound to a cutinase-like enzyme, Cut190*, and its engineering for improved activity. Appl Microbiol Biotech 102: 10067–10077. doi: 10.1007/s00253-018-9374-x
|
| [6] |
Miyakawa T, Mizushima H, Ohtsuka J, et al. (2015) Structural basis for the Ca2+-enhanced thermostability and activity of PET-degrading cutinase from Saccharomonospora viridis AHK190. Appl Microbiol Biotech 99: 4297–4307. doi: 10.1007/s00253-014-6272-8
|
| [7] |
Kawabata T, Oda M, Kawai F (2017) Mutational analysis of cutinase-like enzyme, Cut190, based on the 3D docking structure with model compounds of polyethylene terephthalate. J Biosci Bioeng 124: 28–35. doi: 10.1016/j.jbiosc.2017.02.007
|
| [8] | Inaba S, Kamiya N, Bekker GJ, et al. (2018) Folding thermodynamics of PET-hydrolyzing enzyme Cut190 depending on Ca2+ concentration. J Therm Anal Calorim. |
| [9] |
Numoto N, Kamiya N, Bekker GJ, et al. (2018) Structural dynamics of the PET-degrading cutinase-like enzyme from Saccharomonospora viridis AHK190 in substrate-bound states elucidates the Ca2+ driven catalytic cycle. Biochemistry 57: 5289–5300. doi: 10.1021/acs.biochem.8b00624
|
| [10] |
Masson P, Tonello C, Balny C (2001) High-pressure biotechnology in medicine and pharmaceutical science. J Biomed Biotechnol 1: 85–88. doi: 10.1155/S1110724301000158
|
| [11] |
Buckow R, Weiss U, Heinz V, et al. (2007) Stability and catalytic activity of -amylase from barley malt at different pressure-temperature conditions. Biotechnol Bioeng 97: 1–11. doi: 10.1002/bit.21209
|
| [12] |
Eisenmenger MJ, Reyes-De-Corcuera JI (2009) High pressure enhancement of enzymes: A review. Enzyme Microb Technol 45: 331–347. doi: 10.1016/j.enzmictec.2009.08.001
|
| [13] |
Bamdad F, Bark S, Kwon CH, et al. (2017) Anti-inflammatory and antioxidant properties of peptides released from β-lactoglobulin by high hydrostatic pressure-assisted enzymatic hydrolysis. Molecules 22: E949. doi: 10.3390/molecules22060949
|
| [14] |
Quartinello F, Vajnhandl S, Volmajer Valh J, et al. (2017) Synergistic chemo-enzymatic hydrolysis of poly(ethylene terephthalate) from textile waste. Microb Biotechnol 10: 1376–1383. doi: 10.1111/1751-7915.12734
|
| [15] |
Yamanis J, Vilenchich R, Adelman M (1975) Gas-liquid chromatography of silylated glycols and terephthalate esters. J Chromatogr A 108: 79–84. doi: 10.1016/S0021-9673(00)97490-9
|
| [16] |
Dzwolak W, Kato M, Taniguchi Y (2002) Fourier transform infrared spectroscopy in high-pressure studies on proteins. Biochim Biophys Acta 1595: 131–144. doi: 10.1016/S0167-4838(01)00340-5
|
| [17] |
Imamura H, Isogai Y, Kato M (2012) Differences in the structural stability and cooperativity between monomeric variants of natural and de novo Cro proteins revealed by high-pressure Fourier transform infrared spectroscopy. Biochemistry 51: 3539–3546. doi: 10.1021/bi2019223
|
| [18] |
Wong PTT, Moffatt D (1989) A new internal pressure calibrant for high-pressure infrared spectroscopy of aqueous systems. Appl Spectrosc 43: 1279–1281. doi: 10.1366/0003702894203642
|
| [19] |
Savitzky A, Golay MJE (1964) Smoothing and differentiation of data by simplified least squares procedures. Anal Chem 36: 1627–1639. doi: 10.1021/ac60214a047
|
| [20] |
Barth A, Zscherp C (2002) What vibrations tell us about proteins. Q Rev Biophys 35: 369–430. doi: 10.1017/S0033583502003815
|
| [21] |
Byler DM, Susi H (1986) Examination of the secondary structure of proteins by deconvolved FTIR spectra. Biopolymers 25: 469–487. doi: 10.1002/bip.360250307
|
| [22] |
Torii H, Tasumi M (1992) Model calculations on the amide-I infrared bands of globular proteins. J Chem Phys 96: 3379–3387. doi: 10.1063/1.461939
|
| [23] |
Ganim Z, Chung HS, Smith AW, et al. (2008) Amide I two-dimensional infrared spectroscopy of proteins. Acc Chem Res 41: 432–441. doi: 10.1021/ar700188n
|
| [24] |
Koschorreck K, Liu D, Kazenwadel C, et al. (2010) Heterologous expression, characterization and site-directed mutagenesis of cutinase CUTAB1 from Alternaria brassicicola. Appl Microbiol Biotechnol 87: 991–997. doi: 10.1007/s00253-010-2533-3
|
| [25] |
Chen S, Tong X, Woodard RW, et al. (2008) Identification and characterization of bacterial cutinase. J Biol Chem 283: 25854–25862. doi: 10.1074/jbc.M800848200
|
| [26] |
Kwon MA, Kim HS, Yang TH, et al. (2009) High-level expression and characterization of Fusarium solani cutinase in Pichia pastoris. Protein Expression Purif 68: 104–109. doi: 10.1016/j.pep.2009.06.021
|
| [27] |
Kim HR, Song WS (2010) Optimization of papain treatment for improving the hydrophilicity of polyester fabrics. Fibers Polym 11: 67–71. doi: 10.1007/s12221-010-0067-z
|
| [28] |
Billig S, Oeser T, Birkemeyer C, et al. (2010) Hydrolysis of cyclic poly(ethylene terephthalate) trimers by a carboxylesterase from Thermobifida fusca KW3. Appl Microbiol Biotechnol 87: 1753–1764. doi: 10.1007/s00253-010-2635-y
|
| [29] |
Barth M, Oeser T, Wei R, et al. (2015) Effect of hydrolysis products on the enzymatic degradation of polyethylene terephthalate nanoparticles by a polyester hydrolase from Thermobifida fusca. Biochem Eng J 93: 222–228. doi: 10.1016/j.bej.2014.10.012
|
| [30] |
Joo S, Cho IJ, Seo H, et al. (2018) Structural insight into molecular mechanism of poly(ethylene terephthalate) degradation. Nat Commun 9: 382. doi: 10.1038/s41467-018-02881-1
|
| [31] |
Gall A, Ellervee A, Sturgis JN, et al. (2003) Membrane protein stability: high pressure effects on the structure and chromophore-binding properties of the light-harvesting complex LH2. Biochemistry 42: 13019–13026. doi: 10.1021/bi0350351
|
| [32] |
Dirix C, Duvetter T, Loey AV, et al. (2005) The in situ observation of the temperature and pressure stability of recombinant Aspergillus aculeatus pectin methylesterase with Fourier transform IR spectroscopy reveals an unusual pressure stability of β-helices. Biochem J 392: 565–571. doi: 10.1042/BJ20050721
|
 biophy-05-04-290-s001.pdf biophy-05-04-290-s001.pdf |
 |
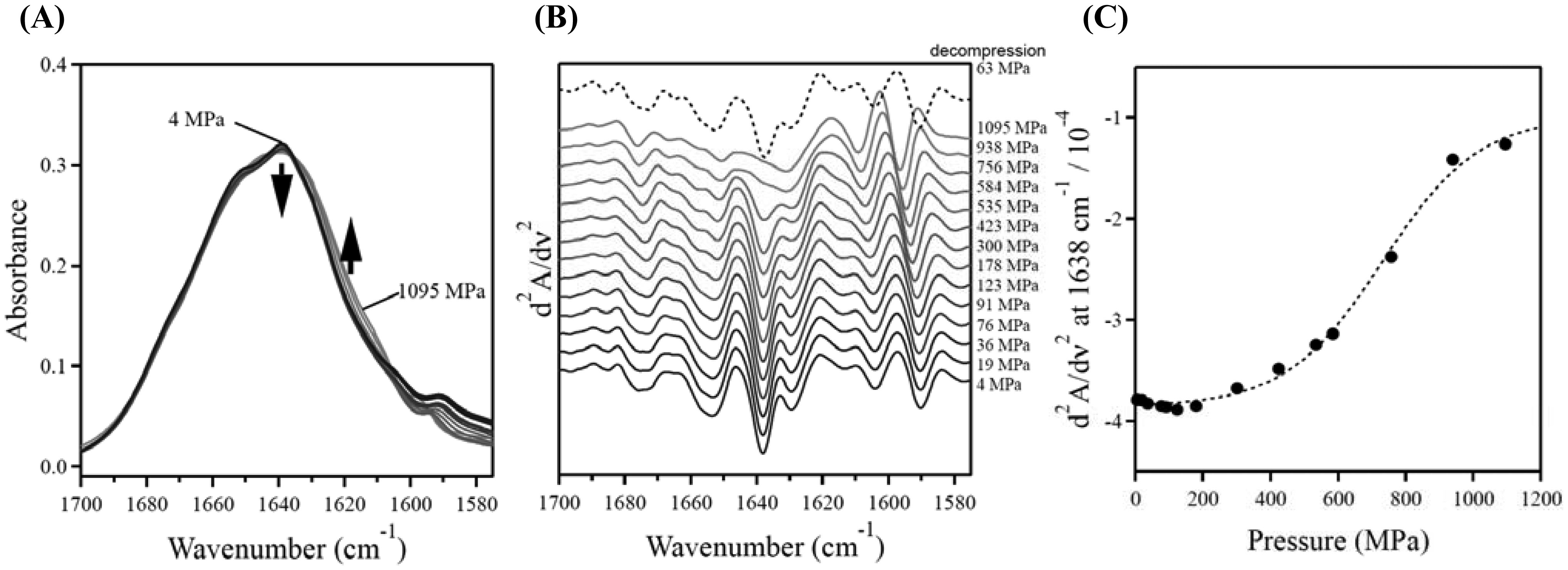
Figures(5)

Yoshiji Hantani, Hiroshi Imamura, Tsubasa Yamamoto, Akane Senga, Yuri Yamagami, Minoru Kato, Fusako Kawai, Masayuki Oda. Functional characterizations of polyethylene terephthalate-degrading cutinase-like enzyme Cut190 mutants using bis(2-hydroxyethyl) terephthalate as the model substrate[J]. AIMS Biophysics, 2018, 5(4): 290-302. doi: 10.3934/biophy.2018.4.290







 DownLoad:
DownLoad: