Bacterial lysates could alleviate the clinical symptoms of allergic rhinitis (AR) and decrease the recurrent rate of AR through regulation of regulatory T cell (Treg) cells. However, the molecular regulatory mechanisms of bacterial lysates to Treg are still unclear.
We aimed to investigate the importance of microRNA-155 (miR-155) to Treg cells function in OM-85 Broncho-Vaxom (OM-85 BV) treated experimental mouse models of AR.
AR mouse models were established and treated by intranasal administration of OM-85 BV to investigate the role of bacteria lysate for Treg cell function. The proliferation of Treg cells in peripheral blood was examined. The mRNA levels of IL-10, transforming growth factor-β (TGF-β) were examined by real-time PCR. miR-155 mimics and inhibitor were used to verify the role of miR-155 for Treg cells function.
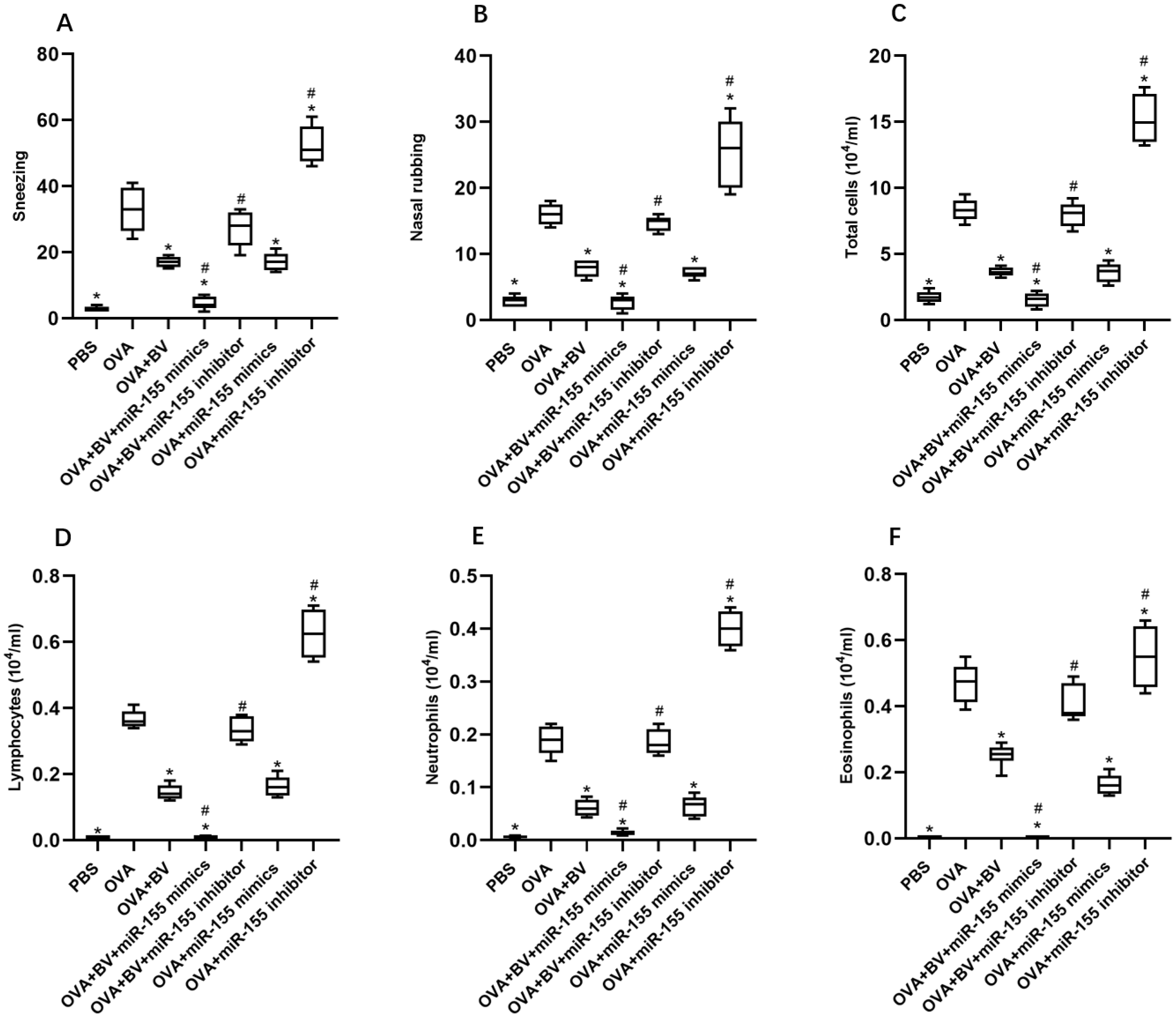
OM-85 BV, miR-155 mimics or their combination reduced total cells, lymphocytes, neutrophils and eosinophils in nasal lavage fluid of AR mouse models and improved allergic symptoms. OM-85 BV promoted the proliferation of Treg and the expression of Foxp3, IL-10 and TGF-β both in vivo and in vitro. The miR-155 enhanced the proliferation and function of Treg.
MiR-155 promotes Treg cells function in OM-85 BV bacteria lysate treated experimental models of AR and alleviate the upper airway allergic inflammation in AR mice.
Citation: Xi Luo, Hehong Li, Rongshan Chen, Yinhui Zeng, Wenlong Liu, Qingxiang Zeng. MicroRNA-155 is a critical regulator of regulatory T cells in OM-85 Broncho-Vaxom treated experimental models of allergic rhinitis[J]. AIMS Allergy and Immunology, 2024, 8(1): 8-17. doi: 10.3934/Allergy.2024002
Bacterial lysates could alleviate the clinical symptoms of allergic rhinitis (AR) and decrease the recurrent rate of AR through regulation of regulatory T cell (Treg) cells. However, the molecular regulatory mechanisms of bacterial lysates to Treg are still unclear.
We aimed to investigate the importance of microRNA-155 (miR-155) to Treg cells function in OM-85 Broncho-Vaxom (OM-85 BV) treated experimental mouse models of AR.
AR mouse models were established and treated by intranasal administration of OM-85 BV to investigate the role of bacteria lysate for Treg cell function. The proliferation of Treg cells in peripheral blood was examined. The mRNA levels of IL-10, transforming growth factor-β (TGF-β) were examined by real-time PCR. miR-155 mimics and inhibitor were used to verify the role of miR-155 for Treg cells function.
OM-85 BV, miR-155 mimics or their combination reduced total cells, lymphocytes, neutrophils and eosinophils in nasal lavage fluid of AR mouse models and improved allergic symptoms. OM-85 BV promoted the proliferation of Treg and the expression of Foxp3, IL-10 and TGF-β both in vivo and in vitro. The miR-155 enhanced the proliferation and function of Treg.
MiR-155 promotes Treg cells function in OM-85 BV bacteria lysate treated experimental models of AR and alleviate the upper airway allergic inflammation in AR mice.

| [1] |
Genuneit J, Standl M (2022) Epidemiology of allergy: Natural course and risk factors of allergic diseases. Handb Exp Pharmacol 268: 21-27. https://doi.org/10.1007/164_2021_507 
|
| [2] |
Li F, Zhou Y, Li S, et al. (2011) Prevalence and risk factors of childhood allergic diseases in eight metropolitan cities in China: a multicenter study. BMC Public Health 11: 437. https://doi.org/10.1186/1471-2458-11-437
|
| [3] |
Barnes PJ (2011) Pathophysiology of allergic inflammation. Immunol Rev 242: 31-50. https://doi.org/10.1111/j.1600-065X.2011.01020.x
|
| [4] |
Zhang Y, Lan F, Zhang L (2021) Advances and highlights in allergic rhinitis. Allergy 76: 3383-3389. https://doi.org/10.1111/all.15044
|
| [5] |
Tomazic PV, Lang-Loidolt D (2021) Current and emerging pharmacotherapy for pediatric allergic rhinitis. Expert Opin Pharmacother 22: 849-855. https://doi.org/10.1080/14656566.2020.1808622
|
| [6] |
Janeczek K, Kaczyńska A, Emeryk A, et al. (2022) Perspectives for the use of bacterial lysates for the treatment of allergic rhinitis: A systematic review. J Asthma Allergy 15: 839-850. https://doi.org/10.2147/jaa.S360828
|
| [7] |
Kaczynska A, Klosinska M, Janeczek K, et al. (2022) Promising immunomodulatory effects of bacterial lysates in allergic diseases. Front Immunol 13: 907149. https://doi.org/10.3389/fimmu.2022.907149
|
| [8] |
Koatz AM, Coe NA, Cicerán A, et al. (2016) Clinical and immunological benefits of OM-85 bacterial lysate in patients with allergic rhinitis, asthma, and COPD and recurrent respiratory infections. Lung 194: 687-697. https://doi.org/10.1007/s00408-016-9880-5
|
| [9] |
Specjalski K, Jassem E (2019) MicroRNAs: Potential biomarkers and targets of therapy in allergic diseases?. Arch Immunol Ther Exp 67: 213-223. https://doi.org/10.1007/s00005-019-00547-4
|
| [10] | Zhu YQ, Liao B, Liu YH, et al. (2019) MicroRNA-155 plays critical effects on Th2 factors expression and allergic inflammatory response in type-2 innate lymphoid cells in allergic rhinitis. Eur Rev Med Pharmacol Sci 23: 4097-4109. https://doi.org/10.26355/eurrev_201905_17911 |
| [11] |
Johansson K, Malmhäll C, Ramos-Ramírez P, et al. (2017) MicroRNA-155 is a critical regulator of type 2 innate lymphoid cells and IL-33 signaling in experimental models of allergic airway inflammation. J Allergy Clin Immunol 139: 1007-1016. https://doi.org/10.1016/j.jaci.2016.06.035
|
| [12] |
Zeng Q, Liu W, Luo R, et al. (2019) MicroRNA-181a and microRNA-155 are involved in the regulation of the differentiation and function of regulatory T cells in allergic rhinitis children. Pediatr Allergy Immunol 30: 434-442. https://doi.org/10.1111/pai.13038
|
| [13] |
Lee SH, Kim HJ, Lee SY, et al. (2023) Broncho-Vaxom bacterial lysate prevents asthma via acetate enhancement in mouse model. Pediatr Allergy Immunol 34: e14018. https://doi.org/10.1111/pai.14018
|
| [14] |
Fu R, Li J, Zhong H, et al. (2014) Broncho-Vaxom attenuates allergic airway inflammation by restoring GSK3β-related T regulatory cell insufficiency. PLoS One 9: e92912. https://doi.org/10.1371/journal.pone.0092912
|
| [15] |
Shen Y, Li L, Chen W, et al. (2023) Apolipoprotein E negatively regulates allergic airway inflammation and remodeling in mice with OVA-induced chronic asthma. Int Immunopharmacol 116: 109776. https://doi.org/10.1016/j.intimp.2023.109776
|
| [16] |
Lau S, Gerhold K, Zimmermann K, et al. (2012) Oral application of bacterial lysate in infancy decreases the risk of atopic dermatitis in children with 1 atopic parent in a randomized, placebo-controlled trial. J Allergy Clin Immunol 129: 1040-1047. https://doi.org/10.1016/j.jaci.2012.02.005
|
| [17] |
Janeczek KP, Emeryk A, Rapiejko P (2019) Effect of polyvalent bacterial lysate on the clinical course of pollen allergic rhinitis in children. Postepy Dermatol Alergol 36: 504-505. https://doi.org/10.5114/ada.2019.87457
|
| [18] | Meng Q, Li P, Li Y, et al. (2019) Broncho-vaxom alleviates persistent allergic rhinitis in patients by improving Th1/Th2 cytokine balance of nasal mucosa. Rhinology 57: 451-459. https://doi.org/10.4193/Rhin19.161 |
| [19] |
Han L, Zheng CP, Sun YQ, et al. (2014) A bacterial extract of OM-85 Broncho-Vaxom prevents allergic rhinitis in mice. Am J Rhinol Allergy 28: 110-116. https://doi.org/10.2500/ajra.2013.27.4021
|
| [20] |
Kohlhaas S, Garden OA, Scudamore C, et al. (2009) Cutting edge: the Foxp3 target miR-155 contributes to the development of regulatory T cells. J Immunol 182: 2578-2582. https://doi.org/10.4049/jimmunol.0803162
|
| [21] |
Schjenken JE, Moldenhauer LM, Zhang B, et al. (2020) MicroRNA miR-155 is required for expansion of regulatory T cells to mediate robust pregnancy tolerance in mice. Mucosal Immunol 13: 609-625. https://doi.org/10.1038/s41385-020-0255-0
|


Figures(3)

Xi Luo, Hehong Li, Rongshan Chen, Yinhui Zeng, Wenlong Liu, Qingxiang Zeng. MicroRNA-155 is a critical regulator of regulatory T cells in OM-85 Broncho-Vaxom treated experimental models of allergic rhinitis[J]. AIMS Allergy and Immunology, 2024, 8(1): 8-17. doi: 10.3934/Allergy.2024002







 DownLoad:
DownLoad: